|
Glacier
Homing Behavior of Transplanted Black Bears, Glacier National Park Katherine L. McArthur |

|
HOMING BEHAVIOR OF TRANSPLANTED BLACK BEARS,
GLACIER NATIONAL PARK
National Park Service Progress Report
For use by National Park Service and cooperating agencies
Katherine L. McArthur
Research Division
Glacier National Park
West Glacier, Montana 59936
May 30, 1978
HOMING BEHAVIOR OF TRANSPLANTED BLACK BEARS, GLACIER NATIONAL PARK
KATHERINE L. McARTHUR, Glacier National Park, West Glacier, Montana 59936
Abstract: Data on the transplanting of 101 black bears (Ursus americanus) on 155 occasions between 1967 and 1976 in Glacier National Park were evaluated to examine the homing behavior of transplanted black bears and to identify factors that contribute to transplant success. Females were more likely than males to return to the capture area; males were more likely than females to become nuisances in developed areas near the release sites. The more a bear weighed, up to 100 kg, the more likely it was to return to the capture area; two-thirds of the transplants of bears weighing 100 kg or more were successful. Transplant success was very highly correlated with distance of the transplant and with the estimated elevation gain that a homing bear would encounter in returning; presence of a physiographic barrier also contributed to transplant success. It is suggested that male bears that repeatedly exhibit nuisance behavior may have difficulty in obtaining adequate natural forage. Evidence suggested that a portion of the adult population exists as transients, with very large home ranges. Implications for management are discussed.
TABLE OF CONTENTS
Where wilderness occupied by black bears interfaces with human-use areas, bears are commonly attracted into the developed areas by garbage, carelessly stored food, and other unnatural sources of food (Barnes and Bray 1967, Sauer et al. 1969, Piekielek and Burton 1975, Beeman and Pelton 1976, Lindzey et al. 1976, Harms 1978). When a bear enters a developed area for the first time, its actions may be governed largely by chance, but if a behavior pattern is reinforced by a food reward, the bear is likely to repeat it (Stokes 1970:1156). In an effort to obtain food, bears may learn to threaten people or cause property damage or personal injury.
In Glacier National Park, inhabited by an estimated 500 black bears (Glacier National Park 1978), management emphasizes preservation of natural biotic relationships. However, annual visitation increased dramatically from 67,200 in 1945 to 884,000 in 1967 and 1,660,000 in 1976. Associated with increased visitation were rising numbers of bear/human conflicts. Management implemented to resolve these conflicts has involved preventive as well as corrective measures. Elimination of factors that lead to nuisance bear behavior is the most effective and economical method for reducing the number of bear/human conflicts (Beeman and Pelton 1976:87, Harms 1978). Between 1967 and 1970, a concerted effort was begun in Glacier National Park to inform visitors and make unnatural sources of food unavailable to bears. Despite this effort, some unnatural food sources continued to exist, as when bearproof garbage cans were overfilled, when sightseers offered food to bears grazing at the roadside, or when campers failed to securely store their food or properly dispose of garbage.
Corrective management involves capture and transplanting of nuisance bears, and destruction of bears that repeatedly exhibit nuisance behavior. Erickson and Petrides (1964:60) stated that troublesome bears can be transplanted without great likelihood of their return. In contrast, Barnes and Bray (1967:116), Harger (1967:6), Sauer et al. (1969:216-220), Payne (1975:812), Piekielek and Burton (1975:23), and Beeman and Pelton (1976:90) observed that bears have a definite tendency to return to the capture site and that the likelihood that a bear would return was related to the distance it was transplanted. Beeman and Pelton (1976:92) stated that rough topography and dense vegetation may have hindered long movements of bears in the Great Smoky Mountains. No other study has apparently considered topographic features.
I am grateful to C. Martinka for providing the opportunity to conduct this study and for offering valuable guidance and encouragement. I also thank J. DeSanto, M. Meagher, C. Meslow, and J. Peek for their thoughtful and constructive reviews of the manuscript.
STUDY AREA
Glacier National Park, an area of 4,100 km, is situated adjacent to the Canadian border in northwestern Montana. The park is bisected by the Continental Divide; elevations range from 961 m to 3,190 m.
The terrain is very rugged. Physiographic features reflect glacial and, to a lesser extent, fluvial erosion of the uplifted sedimentary rock. Most precipitation falls west of the Continental Divide. Up to 250 cm of precipitation falls at upper elevations, mostly as snow (Glacier National Park 1976:23).
The west side of the park and portions of the east side are blanketed by spruce (Picea engelmanni x glauca) / fir (Abies lasiocarpa) forests and various successional stages of lodgepole pine (Pinus contorta) communities. Bunchgrass (Agropyron spicatum, Festuca spp.) communities are common on the east side of the park and occur locally on the west side. One-third of the park lies above timberline.
In addition to black bears, the park is inhabited by approximately 200 grizzly bears (Ursus arctos) (Martinka 1974:23).
METHODS
Most bears were captured in or near developed areas or along roads using baited culvert traps (Erickson 1957) and subsequently immobilized. A limited number of free-ranging bears were immobilized with dart guns employing succinylcholine chloride (Sucostrin) or etorphine (M99). Each immobilized bear was marked with a numbered metal tag in one or both ears. Sex and weight of the bear were determined.
Bears were transported to remote sites within the park and released. Release sites were selected by considering distance from the capture site and proximity to other developed areas. During transport, bears were not drugged and were able to see out of the culvert trap.
Data analyzed included characteristics of the bear as well as of the transplant. Characteristics of the bear included weight; sex; whether cubs were present and, if present, whether they were also transplanted; and the number of times the bear had been previously transplanted. Since the age of the bear was unknown in most cases, the bear's weight was used as a general indicator of its age. Whether the bear obtained unnatural food at the capture area usually had not been recorded; the influence of this factor therefore could not be evaluated. Characteristics of the transplant included date, straight-line distance, and the number of drainages away that the bear was transplanted. In addition, the route that a homing bear might take was simulated by plotting a straight-line route and adjusting it to consider topographic features such as mountain passes and skirting ridges instead of going directly over them; the total elevation gain that the bear would encounter in returning to the capture area along this route was estimated. Also noted was whether the homing bear would encounter a physiographic barrier such as a sharp ridge at the head of a valley, or be forced by the terrain to travel outside the eastern boundary of the park, where there was severe hunting pressure. Data were not generally available on the number of days until a bear returned, only on the number of days until it was recaptured.
A transplant was considered successful if the transplanted bear was not seen again through 1977. An unsuccessful transplant was defined as one from which the bear returned to the capture area. Other possible outcomes were that (1) the bear became a nuisance in another area, or (2) the bear was killed outside the park.
Data were analyzed using standard statistical analyses including chi-square, t-test, and linear regression. For analysis, transplants were separated into first and subsequent transplants. An experienced bear was defined as a bear that had been previously transplanted.
RESULTS
A total of 101 black bears were transplanted 155 times between 1967 and 1976 (Table 1). Thirty-five bears were successfully transplanted the first time; 18 were successfully transplanted on subsequent transplants.
Table 1. Transplant success associated with various characteristics of transplanted black bears, Glacier National Park, 1967-76. Percentages are in parentheses.
| Characteristic | Successful | Returned to capture area |
Nuisance in different area |
Killed in different area |
Total |
| First-time transplants | 34 (35) | 50 (49) | 11 (11) | 5 (5) | 101 |
| Subsequent transplants | 18 (33) | 27 (50) | 6 (11) | 3 (6) | 54 |
| Total | 53 (34) | 77 (50) | 17 (11) | 8 (5) | 155 |
| Sex | |||||
| Male | 29 (38) | 31 (40) | 14 (18) | 3 (4) | 77 |
| Female | 17 (27) | 40 (63) | 3 (5) | 4 (6) | 64 |
| Unknown | 7 (50) | 6 (43) | 0 (0) | 1 (7) | 14 |
| Cubs present | |||||
| Entire litter transported | 2 (40) | 1 (20) | 2 (40) | 0 (0) | 5 |
| Partial litter transported | 1 (50) | 1 (50) | 0 (0) | 0 (0) | 2 |
| No cubs transported | 0 (0) | 3 (100) | 0 (0) | 0 (0) | 3 |
| Weight (kg) | |||||
| 0—19 | 1 (17) | 1 (17) | 3 (50) | 1 (17) | 6 |
| 20—39 | 11 (42) | 9 (35) | 4 (15) | 2 (8) | 26 |
| 40—59 | 14 (26) | 32 (60) | 6 (11) | 1 (2) | 53 |
| 60—79 | 14 (45) | 16 (52) | 1 (3) | 0 (0) | 31 |
| 80—99 | 1 (13) | 7 (87) | 0 (0) | 0 (0) | 8 |
| >100 | 4 (67) | 1 (17) | 1 (17) | 0 (0) | 6 |
| Unknown | 8 (32) | 11 (44) | 2 (8) | 4 (16) | 25 |
Characteristics of the Bear
Erickson and Petrides (1964:50), Harger (1967:7), Sauer et al. (1969:217), Piekielek and Burton (1975:22), and Beeman and Pelton (1976:90) found that 78-91 percent of nuisance bears were males. In Glacier National Park, sex of 91 percent of the nuisance bears was recorded. Of these, 55 percent were males and 45 percent were females; this ratio is significantly different from those of the previous studies (P<0.05).
Transplants of female bears were somewhat less successful than were transplants of male bears (Table 1). Females were more likely than males to return to the capture area; transplanted males were more likely than females to become nuisances in other areas. This observation supports the statement of Bunnell and Tait (1978:48, 51) that female bears defend specific territories while males do not.
Beeman and Pelton (1976:50) reported that only 1 (1.3 percent) of 76 nuisance bears was a female with cubs; in contrast, 64 (41.3 percent) of 155 transplants of nuisance bears in Glacier involved females, and 10 (15.6 percent) of the 64 females had cubs with them. When at least some of the cubs were transplanted with the mother, the transplants met with varying degrees of success (Table 1). In each. of 3 instances in which a female was transplanted without any of her cubs, the female returned to the capture site. One of these females returned 11 km in 8 hours.
Although Barnes and Bray (1967:118) observed that homing was more prevalent among adults than subadults, Beeman and Pelton (1976:90) found no significant difference between homing of adults and that of subadults. In this study, regression analysis showed that weight of the bear, up to 100 kg, was positively correlated with the percentage of bears that returned to the capture area (Y = 0.785 X + 10.950, r = 0.9405). In other words, the more a bear weighed, up to 100 kg, the more likely it was to return to the capture area. In direct contrast, however, 67 percent of the transplants of bears weighing 100 kg or more were successful; in fact, this was the most successful weight category. The reason for this relationship could not be ascertained.
Although the differences were not significant (P>0.50), female bears successfully transplanted on their first transplant tended to weigh less than those successfully transplanted on a subsequent transplant (Table 2). This pattern also held true for females and males that returned to the capture area: bears transplanted for the first time tended to weigh less than those transplanted a subsequent time. This tendency was expected because the average bear transplanted for the first time is probably younger than the average bear that has been transplanted previously. However, male bears successfully transplanted on their first transplant tended to weigh more than those whose subsequent transplants were successful. Although bears tend to weigh less earlier in the year, the relationship could not be adequately explained by transplant dates; the mean date of successful first-time transplants of male bears was 17 July and the mean date of successful subsequent transplants of male bears was 15 July (Fig. 1). Male bears that weigh less may be more likely to return to the capture area if they roam more widely and are familiar with a greater area than heavier bears. Another possibility is that male bears that weigh less have difficulty in obtaining adequate natural forage, are more dependent on unnatural food, and therefore have greater incentive to return to a known source of unnatural food. This hypothesis is supported by the fact that the lightest-weight bears were the ones most likely to become nuisances in other developed areas (Table 1).
Table 2. Mean weights (kg) and standard deviations (SD) of females, males, and all bears that were transplanted successfully and those that returned to the capture area, comparing first-time transplants with subsequent transplants, Glacier National Park, 1967-76.
| Female (±SD) |
Male (±SD) | Overalla (±SD) | ||||
| Successful | Returned | Successful | Returned | Successful | Returned | |
| First-time transplants | 47.2 ± 17.2 (N= 10) |
50.8 ± 18.4 (N= 22) |
63.2 ± 27.6 (N= 21) |
59.4 ± 21.1 (N= 22) |
58.7 ± 25.4 (N= 35) |
55.6 ± 19.8 (N= 50) |
| Subsequent transplants | 53.8 ± 20.3 (N= 7) |
51.9 ± 15.0 (N= 18) |
50.0 ± 23.3 (N= 8) |
64.5 ± 15.1 (N= 9) |
51.9 ± 20.6 (N= 18) |
57.0 ± 15.9 (N= 27) |
| Total | 49.6 ± 17.9 (N= 17) |
51.2 ± 17.1 (N= 40) |
59.6 ± 26.7 (N= 29) |
60.8 ± 19.6 (N= 31) |
56.6 ± 24.0 (N= 53) |
56.0 ± 18.6 (N= 77) |
aIncludes bears of unknown sex. | ||||||
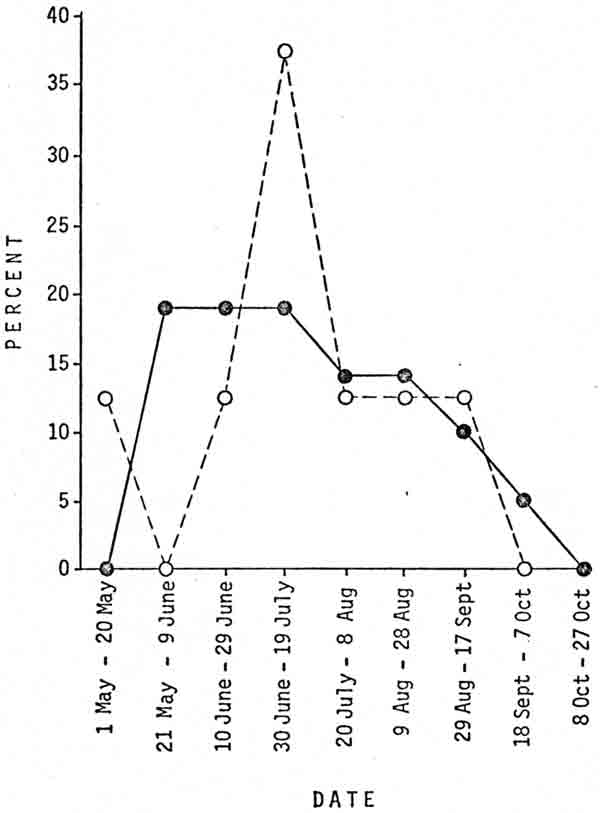
|
| Fig. 1. percentages of successful first-time (—)(N=21) and successful subsequent (— — —)(N=8) transplants of male black bears that took place during 20-day time periods, Glacier National Park, 1967-76. |
The percentage of transplants that were successful was essentially the same between first and subsequent transplants (Table 1). Experienced bears that returned to the capture area had been previously transplanted a mean of 1.7 times, whereas experienced bears that were successfully transplanted had been previously transplanted a mean of 1.4 times; the difference was not significant (P<0.25).
Characteristics of the Transplant
Although increased movements by bears during autumn have been noted (Erickson and Petrides 1964:56, Piekielek and Burton 1975:16), and Barnes and Bray (1967:118) reported greater success of transplants in autumn—both associated with more abundant local food sources—little correlation was found in this study between success and time of year (Y = 0.096 X + 54.569, r = —0.3274), nor was the percentage of bears that returned to the capture area correlated with time of year (Y = 0.012 X + 53.339, r = —0.0724).
Bears transplanted a greater distance were significantly less likely to return to the capture area (Fig. 2). In first-time transplants, the mean distance of successful transplants was 29.9 km, whereas the mean distance of transplants where the bear returned was 19.2 km; the difference was significant (P<0.001). In subsequent transplants, the mean distance of successful transplants was 35.3 km, whereas the mean distance of transplants where the bear returned was 23.6 km; the difference was not significant (P<0.25).
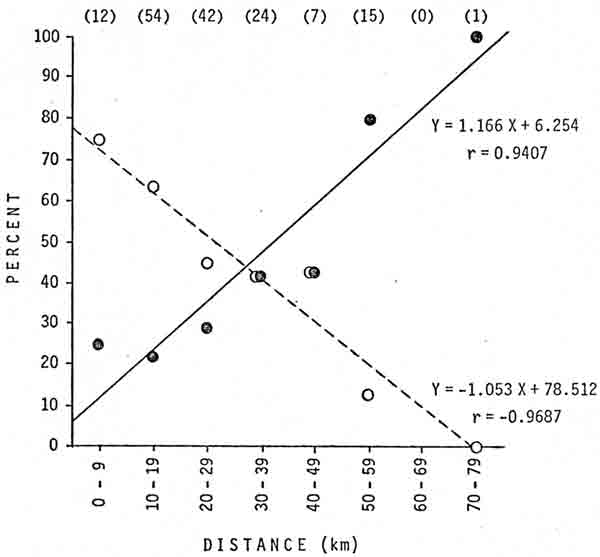
|
| Fig. 2. Percentages of transplants that were successful (—) or from which the bear returned (— — —) in relation to distance of the transplant, Glacier National Park, 1967-76. Sample sizes are in parentheses. |
The number of drainages across which a transplanted bear had to return was also important (Fig. 3). In first-time transplants, the mean number of drainages was 3.3 in successful transplants and 1.6 in transplants where the bear returned; the difference was significant (P<0.01). In subsequent transplants, the mean number of drainages was 4.1 in successful transplants and 2.4 in transplants where the bear returned; the difference was not significant (P<0.25).
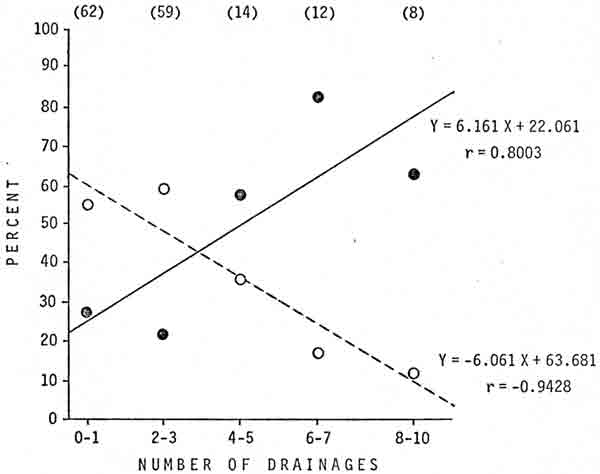
|
| Fig. 3. Percentages of transplants that were successful (—) or from which the bear returned (— — —) in relation to the number of drainages across which the bear was transplanted, Glacier National Park, 1967-76. Sample sizes are in parentheses. |
The elevation gain a bear would encounter in homing from the transplant site to the capture area has not been quantitatively considered in previous studies. This factor was indicated to be an important component of transplant success (Fig. 4). Data for elevations of 1,500-1,999 m were not included in Fig. 4. The sample for those elevations consisted of 3 transplants; none were successful. Two bears returned to the capture area; one took 3 years and one took 7 years to do so. The third bear became a nuisance in another area 2 years later. If these data are included, the regression of percent success on elevation becomes Y = 0.024 X + 17.292, r = 0.7532; and the regression of percent returned to the capture area on elevation becomes Y = —0.017 X + 57.839, r = —0.7386.
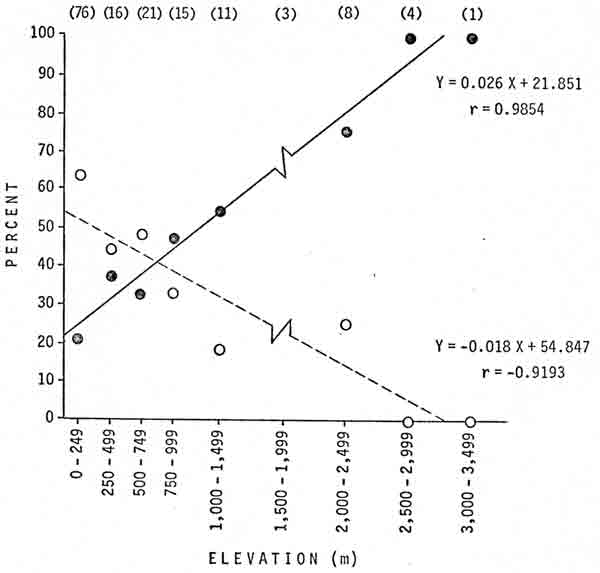
|
| Fig. 4. Percentages of transplants that were successful (—) or from which the bear returned (— — —) in relation to the estimated elevation gain that the homing bear would encounter, Glacier National Park, 1967-76. Sample sizes are in parentheses. |
In first-time transplants, the mean elevation gain involved was 834 m in successful transplants and 352 m in transplants where the bear returned; the difference was significant (P<0.01). In subsequent transplants, the mean elevation gain involved was 1,214 m in successful transplants and 477 m in transplants where the bear returned; again, the difference was significant (P<0.01).
Presence of a physiographic barrier was another apparently important contributor to transplant success. Of 23 transplants where the homing bear would encounter a barrier, 16 (70 percent) were successful; 3 (13 percent) bears returned to the capture area. Three (13 percent) bears become nuisances in other developed areas, indicating that a barrier did not simply shunt a problem bear to another area.
DISCUSSION
Because home range fulfills physical needs as well as psychic needs by allowing bears to move about familiar areas with assurance and to efficiently utilize the resources there, transplanted bears are highly motivated to return to their home ranges (Sauer et al. 1969:216; Beeman and Pelton 1976:93). To preclude bears from returning to the capture area, they must be removed from the area with which they are familiar (Sauer et al. 1969:222). Bears released in unfamiliar surroundings evidently wander at random until they either find a familiar area or can establish new home ranges (Erickson and Petrides 1964:60).
Previous studies have found mean summer home range diameters of female black bears to be 0.9-11.9 km, and those of male black bears to be 2.8-13.8 km (Erickson and Petrides 1964:56, Sauer et al. 1969:212-216, Jonkel and Cowan 1971:19, Poelker and Hartwell 1973:62, Piekielek and Burtin 1975:16). Although these home range studies have found that male bears are wider-ranging than females, females in this study were more apt than males to return to the capture site. Males, on the other hand, were more likely than females to become nuisances in developed areas near the release sites. Evidence presented in this paper indicates that males that repeatedly exhibit nuisance behavior may have difficulty in obtaining adequate natural forage. Poelker and Hartwell (1973:61-62) theorized that bear damage to conifers was related to a shortage of preferred food and that wider ranging of bears in tree-damage areas reflected more extensive searching for food. The statement of Lindzey et al. (1976:219) that nuisance bears range farther than non-nuisance bears is supported by the mean distances from which nuisance bears returned in this study—19.4 km for males and 21.7 km for females—which are substantially greater than reported normal home range sizes.
Several studies have noted unexplained long-distance movements by some bears. Amstrup and Beecham (1976:345) observed a female with a home range of 130.3 km2. Although Erickson and Petrides (1964:57) maintained that bears could generally be transplanted without returning, they reported one bear that returned from a transplant of 103 km and another that returned 154 km. Erickson and Petrides (1964:56) also reported that a female with a cub moved 31.2 km, while summer movements of other females averaged 2.3 km. Jonkel and Cowan (1971:19) noted unusually long movements by 5 adult bears. Such movements have generally been designated as excursions outside of a smaller home range, and are undoubtedly important in familiarizing a bear with terrain and resources beyond its normal home range and assist it in orienting itself after being transplanted from its home range (Beeman and Pelton 1976:93). On the other hand, as suggested by Lindzey and Meslow (1977:421), these may be transient adults, similar to mountain lions (Felis concolor) described by Seidensticker et al. (1973:55) and to adult "floaters" reported in several bird species (Smith 1978). A mixture of resident and transient adults would promote more complete utilization of resources and ensure the availability of a breeding-age replacement should a resident bear be removed from the population. Transient adults would not receive the benefits of intimate knowledge of a relatively small home range, but would be familiar with a large area and able to take advantage of widely dispersed sources of locally abundant food. Such individuals would be difficult to transplant successfully. They also may make up a large proportion of the nuisance bear population.
MANAGEMENT IMPLICATIONS
Transplanting is an effective method of dealing with nuisance bears, as indicated by the fact that 53 of 101 bears in this study were eventually successfully transplanted. Selection of release sites is an important consideration in the handling of nuisance bears (Barnes and Bray 1967:118, Beeman and Pelton 1976:95). Results of this and other studies show that bears transplanted greater distances are less likely to return to the capture area. Bears should also be transplanted across rough terrain, with large elevation changes and physiographic barriers. Greater effort must be made in transplanting transient bears, which are probably more difficult to successfully transplant because they are familiar with a much larger area than are resident bears.
An abundance of natural food at the release site may be a major factor in determining the success of a transplant (Piekielek and Burton 1975:23-24). Sauer et al. (1969:222) stated that where sources of unnatural food are available near the release site, transplanting a bear will simply transfer the problem. This study showed that females and lighter-weight male bears were especially likely to return to the capture area or to become nuisances in developed areas near the release site. This may be due to the relative lack of easily obtainable food at the release site. More research needs to be done concerning the influence of the abundance and quality of food at release sites.
Additional research should also be done to ascertain the activities and fates of transplanted bears that are not seen again—bears that, in this study, were considered successfully transplanted. Results of studies on the responses of bears released in unfamiliar surroundings will be useful in programs to reestablish populations of threatened or endangered species in formerly occupied habitat.
LITERATURE CITED
Amstrup, S. C., and J. Beecham. 1976. Activity patterns of radio-collared black bears in Idaho. J. Wildl. Manage. 40(2):340-348.
Barnes, V. G., Jr., and O. E. Bray. 1967. Population characteristics and activities of black bears in Yellowstone National Park: final report. Colorado Cooperative Wildlife Research Unit, Colorado State Univ., Fort Collins. 199pp.
Beeman, L. E., and M. R. Pelton. 1976. Homing of black bears in the Great Smoky Mountains National Park. Pages 87-95 in M. R. Pelton, J. W. Lentfer, and G. E. Folk, Jr., eds. Bears—their biology and management. IUCN Publ. New Ser. 40.
Bunnell, F. L., and D. E. N. Tait. 1978. Population dynamics of bears and their implications. Proc. Int. Conf. on Population Dynamics of Large Mammals, Logan, Utah. 64pp. (In press.)
Erickson, A. W. 1957. Techniques for live-trapping and handling black bears. Trans. N. Am. Wildl. Conf. 22:520-543.
_________, and G. A. Petrides. 1964. Population structure, movements, and mortality of tagged bears in Michigan. Pages 46-67 in A. W. Erickson, J. Nellor, and G. A. Petrides. The black bear in Michigan. Michigan Agric. Exp. Stn. Res. Bull. 4.
Glacier National Park. 1976. Final environmental statement: master plan. USDI National Park Service, Washington, D. C. 74pp.
_________. 1978. Bear management plan. USDI National Park Service, West Glacier, Montana. 20pp. + app.
Harger, E. M. 1967. Homing behavior of black bears. Michigan Dept. Conserv. Res. and Devel. Rep. 118. 13pp.
Harms, D. R. 1978. Black bear management in Yosemite National Park. In C. J. Martinka, ed. Bears—their biology and management. Proc. Int. Conf. on Bear Research and Management 4. (In press.)
Jonkel, C. J., and I. McT. Cowan. 1971. The black bear in the spruce-fir forest. Wildl. Monogr. 27. 57pp.
Lindzey, F. G., and E. C. Meslow. 1977. Home range and habitat use by black bears in southwestern Washington. J. Wildl. Manage. 41(3):413-425.
Lindzey, J. S., W. S. Kordek, G. J. Matula, Jr., and W. P. Piekielek. 1976. The black bear in Pennsylvania—status, movements, values, and management. Pages 215-224 in M. R. Pelton, J. W. Lentfer, and G. E. Folk, Jr., eds. Bears—their biology and management. IUCN Publ. New Ser. 40.
Martinka, C. J. 1974. Population characteristics of grizzly bears in Glacier National Park, Montana. J. Mammal. 55(1):21-29.
Payne, N. F. 1975. Unusual movements of Newfoundland black bears. J. Wildl. Manage. 39(4):812-813.
Piekielek, W., and T. S. Burton. 1975. A black bear population study in northern California. California Fish and Game 61(1):4-25.
Poelker, R. J., and H. D. Hartwell. 1973. Black bear of Washington. Washington State Game Dept. Biol. Bull. 14. 180pp.
Seidensticker, J. C., IV, M. G. Hornocker, W. V. Wiles, and J. P. Messick. 1973. Mountain lion social organization in the Idaho Primitive Area. Wildl. Monogr. 35. 60pp.
Smith, S. M. 1978. The "underworld" in a territorial sparrow: adaptive strategy for floaters. Am. Nat. 112(985):571-582.
Stokes, A. W. 1970. An ethologist's views on managing grizzly bears. BioScience 20(21):1154-1157.
mcarthur1/index.htm
Last Updated: 8-Apr-2013