|
Mount Rainier
Environment, Prehistory & Archaeology of Mount Rainier National Park, Washington |

|
Chapter 1:
ENVIRONMENT AND LAND-USE ON MOUNT RAINIER & IN THE SOUTHERN WASHINGTON CASCADES
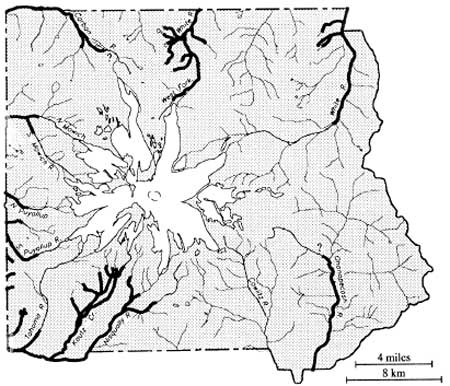
At 14,410 ft, Mount Rainier is the highest, most massive peak in the Cascade Range. It is surrounded by a relatively broad massif of lesser, though locally precipitous mountains, most in the range of 6,000 to 7,000 ft in elevation. Because of combined effects of elevation, mass, latitude and position relative to Pacific westerlies, Mount Rainier sustains the single largest glacier system in the 48 contiguous states. These glaciers are the source of several major Northwest rivers–the Nisqually, Mowich/Puyallup, Carbon and White Rivers draining into Puget Sound; and the Ohanapecosh/Cowlitz system emptying into the Columbia River at Longview north of Portland (Harris 1988:231). Figure 1.1 on page 2 shows a simplified projection of the mountain and its relationship to surrounding landforms and rivers.
Park boundaries form an approximately square 235,612 acre box around the base of the mountain and the northern fringe of the Tatoosh Range on Rainier's southern flank. Environmental characteristics vary with elevation, relative moisture (that is, lee versus windward flanks) and landform. Over the years, different taxonomic systems have been employed to characterize environmental patterns across Mount Rainier and its surrounding terrain (e.g., Brockman's [1947] use of elevational life zones, Franklin and Dyrness' [1973] classic vegetation zones of Oregon and Washington, and Moir's [1989] matrix treatment of Mount Rainier forests). Such schemes emphasize gross patterns in floral, principally forest, composition to present a complex reality in a simpler more comprehensible form.
In this chapter, Mount Rainier's environmental characteristics are classified in a five-part system that draws on existing environmental models–particularly Franklin and Dyrness (1973), but does so in a manner that emphasizes vegetation/resource zones of differential value to prehistoric human populations. Zones used here include 1) expansive low to mid-elevation Northwest maritime forests; 2) major river systems and associated floodplains; 3) subalpine parklands; 4) alpine tundra; and 5) perpetual snowfields, glaciers and glacial scree slopes. While not a vegetation zone per se, the rivers and floodplains distinction accommodates resources (especially anadromous fish) and land-use practices not otherwise expected in the forest to alpine environment. The system is similar to that employed by the author to examine archaeological/environmental associations on the flanks of Mt. Hood in the northern Oregon Cascades (Burtchard and Keeler 1991), and is compatible with systems used to characterize Olympic National Park (Schalk 1988) and North Cascades National Park (Mierendorf 1986) environments. With the addition of lower elevation or more xeric associations, the system can readily be extended to other landforms in and adjacent to the Cascades.
Use of an environmental zone model to characterize Holocene floral and faunal associations (and human land-use patterns) is complicated by at least two variables–dynamic Mount Rainier geology, and Holocene climate change. To gain some control over these issues, this chapter is organized into sections that describe the nature and archaeological implications of each. The first section, is a discussion of local geology with general implications for environment, land-use and the archaeological record. The second describes basic environmental/resource patterns, emphasizing resources believed to be particularly important for attracting systematic human use during prehistoric times. The third section discusses the possible impact of Holocene environmental changes on land-use patterns, and suggests means to improve our understanding of regional paleoclimate. A summary of Mount Rainier environment and long-term use by human populations concludes the chapter.
MOUNT RAINIER AND SOUTHERN WASHINGTON CASCADES
GEOLOGY
The Cascade Range may be divided into two north to south physiographic provinces with Mount Rainier in the middle. The broad North Cascades province extends from the Frazier River in southern British Columbia to Snoqualamie Pass between Glacier Peak and Mount Rainier. The more elongate, narrower South Cascades province extends from the pass (or Mount Rainier) south, across the Columbia River, ultimately terminating at California's Mount Lassen (see Harris 1988 and Whitney 1983:15-30; Burtchard 1990:33-36). Partially due to its position at the northern extreme of the southern province, Mount Rainier's geology shares certain characteristics of each.
Environmental Implications of General Cascades
Geology
There are substantial differences between North and South Cascade physiographic provinces that influence environmental and resource patterns in each. [4] The North Cascades Province is a massive area of old metamorphic and sedimentary rock rafted ashore over 50 million years ago. In origin, these mountains are more closely related to British Columbia's Coast Mountains and Washington's Olympics than to the Cascades further south. The North Cascades also are a substantially broader and higher massif composed of rugged high elevation peaks with deeply incised valleys. Vertical relief tends to be high with elevations ranging from near sea level in major valleys to over 9,000 ft on many of the North Cascade peaks. Due to elevation, breadth, and proximity to moist Pacific storms, the province supports the largest overall number of active glaciers in the lower 48 states.
North Cascades geomorphology has had a marked influence on regional climate, exploitable resource structure and, hence, human use throughout the Holocene. Regional rock types include a variety of geologically distinct exposures, a number of which are suitable for tool manufacture. Forest cover is influenced by exposure and elevation. Maritime forests blanket the western slopes from sea level to timberline at about 5,500 ft. On the eastern slope, forests extend from about 1,000 ft to timberline at 6,500 ft. Timberline on both sides is controlled by deep snow pack that persists into midsummer. It is possible that presence of expansive open habitat supported abundance and variety of floral and faunal resources throughout the Holocene. As in the south, rivers and tributary systems provide entry for anadromous fish. There is little doubt that available resources were sufficient for seasonal use by local populations, provided strategies could accommodate the short growing season and logistical difficulties imposed by rugged terrain, relatively massive breadth and unstable high elevation weather. While not part of the North Cascades per se, it is important to note that most of these environmental qualities and land-use implications apply to Mount Rainier as well.
The South Cascades Province is a generally narrower band of old, highly weathered volcanic mountains, tilted and partially overlain by younger and higher, spatially isolated stratovolcanos along its eastern axis (e.g., Rainier, Adams, Hood, Jefferson, McLoughlin, Shasta and Lassen among others). Most high Cascade mountain building volcanism took place in the Pliocene and Pleistocene 4-1/2 to 3 million years ago, with localized, but significant, activity throughout the Holocene. Except for the high peaks, few of the South Cascade mountains rise above timberline. Forest cover tends to be dense on the western slope grading to more open fir and pine forests on mid to lower elevation eastern slopes. The high peaks, however, are ringed by subalpine and alpine zones that support a variety of floral and faunal resources. Unlike the north, however, parkland and tundra zones are seldom interconnected, but tend to occur as independent, island-like patches within a more uniform forest cover. [5] The South Cascades also have complex drainage systems, many of which support anadromous fish populations.
At a very general level, the entire Cascade Range offered roughly similar economic opportunities and posed comparable constraints to prehistoric people. Both northern and southern provinces offered seasonally available food, fiber, and lithic resources. Severe weather effectively precluded over winter residence in both, and montane landscapes posed varying degrees of logistical difficulty. However, because of their narrower breadth and gentler physiography, the South Cascades probably posed less severe impediments to seasonal use. Vertical relief and distances were less imposing; and with the exception of the highest Cascade peaks, weather was more predictable; making possible longer seasonal use. These effects were counterbalanced to an unknown extent by the small size and isolated character of productive subalpine and alpine resource zones.
Situated near the boundary of the two provinces, Mount Rainier shares characteristics of both. Indeed it is reasonable to refer to montane landscapes from Rainier south to the Columbia River as the southern Washington Cascades physiographic region. Doing so recognizes the area's distinct geological and environmental qualities that distinguish it from its Cascade neighbors to the north and south. Franklin and Dyrness (1973:21-23) offer perhaps the best general summary of these characteristics. Near Mount Rainier, they emphasize the North Cascades-like pattern of accordant ridge crests separated by steep, deeply dissected valleys. Here, ridge crests are relatively high (averaging circa 2,000 m [6,560 ft]), decreasing abruptly to the south (circa 1,200 m [3,936 ft]). Volcanic rock dominates lithic exposures. To a greater extent than elsewhere, southern Washington Cascade landforms are mantled with pumice deposits of variable age, origin and thickness (Franklin and Dyrness 1973:22). Most of these deposits are from Holocene eruptions on Mount Rainier and St. Helens– events which periodically buried the material remains of prehistoric populations using these mountains. Figure 2.1 shows the southern Washington Cascades in relation to surrounding physiographic provinces/regions.
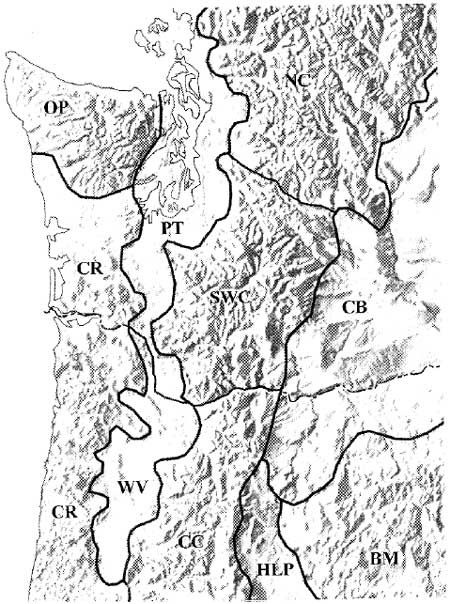
| |||
Figure 2.1 Southern Washington Cascades
and Surrounding Physiographic Provinces (after Rosenfeld 1993:41; see
also Franklin and Dyrness 1973:6. Mount Ranier is about 1/2 inch above
the "C" in SWC)
|
The contrast between the southern Washington Cascades and mountains south of the Columbia River is most sharply visible near Mount Rainier. Mount Rainier's great height and mass supports larger glacial, alpine and subalpine expanses than its southern counterparts. The mountain's size also creates a pronounced rainshadow effect on its northeastern flank, making for more open, patchy forest cover than would otherwise be expected. Furthermore, the breadth, elevation and deeply incised character of the surrounding mountain and ridge system is greater than that further south, mimicking the North Cascades pattern. These qualities make for expansive subalpine and alpine habitats (particularly on Mount Rainier) but make them more logistically challenging to exploit by lowland-based populations. To a greater extent than most of the Southern Cascades, Holocene vulcanism altered the character of the archaeological record through repeated tephra deposits and by destructive mass wasting events (especially lahars).
Mount Rainier Holocene Geology
The modern appearance of Mount Rainier and the southern Washington Cascades is the culmination of Pliocene-Pleistocene mountain building and substantial Holocene modification. By 75,000 years ago, Mount Rainier had reached a height of about 16,000 ft (Harris 1988:240). Though modified by extensive glaciation, the mountain maintained much of that height until after the close of the Fraser/Wisconsin glacial episode circa 15,000 to 13,500 years ago. Most of the subsequent volcanic events that reduced, rebuilt and modified Mount Rainier's shape occurred during the last 7,500 to 5,000 years–a period that undoubtedly witnessed increasing human presence in the general region and on the mountain itself.
The single most massive destructive event (or series of roughly synchronous events) to affect Mount Rainier were the Paradise-Greenwater-Osceola mud and debris flows (lahars) occurring between 5,000 and 4,500 years ago (Scott et al. 1995:12). These events apparently were stimulated by eruption and collapse of Mount Rainier's summit; releasing tremendous volumes of ice, mud and unconsolidated debris down the east and northeastern facies of the mountain (Harris 1988:243). Most of the material (Greenwater and Osceola mudflows) rushed over Steamboat Prow and down both forks of the White River; covering lowland floodplains from the base of the mountain to the Puget lowlands as far away as Auburn, Washington near Tacoma. The Paradise lahar emanated from the same source, but flowed south then west down the Paradise and Nisqually Rivers beyond Ashford, west of the Park boundary. Limited deposits extended down the Nisqually as far as Yelm near Olympia (Crandell 1971; Pringle 1994:2G-1). These events removed about 2,000 vertical feet from Rainier's summit and erased the archaeological record of early Holocene human activity in its affected river valleys. Figure 2.2 shows the extent of the major Mount Rainier lahars, including the more recent Electron mudflow.
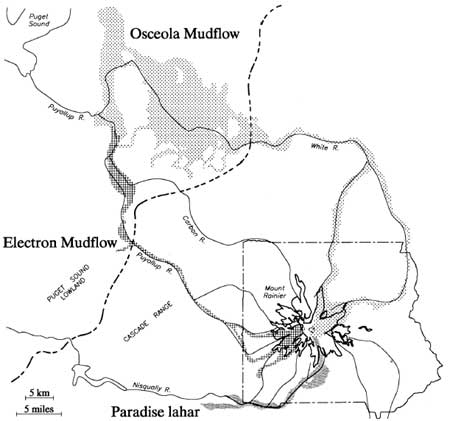
|
| Figure 2.2 Major Mount Rainier Lahars (after Crandell and Mullineaux 1967, and Crandell 1971) |
About 2,500 to 2,300 years ago, renewed eruptive activity on Mount Rainier filled the Osceola crater and rebuilt the mountain summit to approximately its present 14,410 ft height. This eruption ejected coarse-grained tephra (Mount Rainier Pumice Layer C) over the cone and much of the northeast and southeast quadrants of the Park. Gravelly Rainier C tephra is the modern surface on some of the Park's most familiar landscapes such as Sunrise Ridge and rolling alpine tundra between Frozen Lake and Berkeley Park. Deposits range from 2.5 to 20 cm (1 to 8 in) thick. We know that human use of the mountain predates this event by virtue of lithic debris eroding from below Rainier C tephra in borrow pit cut banks on the south slope of Sunrise Ridge (site FS 90-01 or 45PI408). It is likely that more rapid drainage qualities of the gravely tephra contributes to prevalence of patchier forest cover on the mountains lee. Mount Rainier C deposits also make site discovery more difficult, while simultaneously providing a protective, time-diagnostic cap over such deposits once found.
Lava flows associated with the Mount Rainier C eruption were limited to areas near the summit. However, these events spawned lahars which crashed into the White and Nisqually River valleys, again inundating and temporarily raising these valley floors within Park boundaries (Harris 1988:246). On these floodplains, the archaeological record again would have been effectively destroyed.
The most recent major lahar was the Electron mudflow about 500 years ago. This large, clay-rich mudflow (similar in composition to the Osceola flow) poured down the Puyallup and Mowich Rivers on Rainier's western slope. Though less massive than the Osceola flow, it inundated the Puyallup valley floor for almost the same distance into the Puget lowlands (see Figure 2.2). Lesser lahars occurred throughout the Holocene, continuing to the present. The above events are stressed to provide general familiarity with them because of their large scale and consequent impact on the archaeological record. The relatively high frequency and destructive power of such events points to a broader problem in interpreting the archeological record (or absence thereof) along Mount Rainier's river valley floodplains.
Mount Rainier's Holocene geological activity has not been limited to the destructive and creative events noted above. Of some importance to the archaeological record are Holocene tephra producing eruptions by Mount Rainier, Mt. St. Helens, and Mt. Mazama (now Crater Lake) in Oregon. Holocene eruptions from these sources created a complex patchwork of varying depth volcanic deposits, affecting different parts of the Park at different times during the period that humans could have used the mountain. While many of these events provide useful temporal markers for the Park's archaeological record, none are considered to have been of sufficient magnitude to have significantly altered montane land-use patterns in the southern Washington Cascades.
Our understanding of the mountain's volcanic deposition patterns remains incomplete (Pringle pers. com. 1995), but will improve as ongoing dating and mapping studies progress. Available sources have been consulted to provide an outline of depositional eruptive events known to have affected the Park in the last 10,000 years (Table 2.1). Figure 2.3 shows the approximate distribution of the best understood layers. The basic reference for Table 2.1 and Figure 2.3 is Crandell (1987)–who credits most of his information to studies by D.R. Mullineaux. All radiocarbon ages and partial data sets are from Scott et al. (1995) and Pringle (1994).
Table 2.1 Sources and Ages of Volcanic Deposits on Mount Rainier
| Tephra Layer | Thickness (in) | Fragment Dia. (in) | Color | Source | Age (14C years ago) | |
|---|---|---|---|---|---|---|
| West | East | |||||
| X | Absent | 1 | 1/4 - 2 | Light olive gray | Mount Rainier | 100 - 150 |
| W (Wn) | 0 - 1 | 1 - 3 | Medium sand | White | Mt. St. Helens | 470 |
| C | Absent | 1 - 8 | 1/4 - 8 | Brown | Mount Rainier | 2300 |
| P | Mt. St. Helens | 2500 - 3000 | ||||
| Y (Yn) | 5 - 20 | 1 - 5 | Coarse sand | Yellow | Mt. St. Helens | 3400 |
| B | Mount Rainier | 4500 | ||||
| H | Mount Rainier | 4700 | ||||
| F (poss. blast) | Mount Rainier | 5000 | ||||
| S (poss. blast) | Mount Rainier | 5200 | ||||
| D | Absent | 0 - 6 | 1/4 - 6 | Brown | Mount Rainier | 5500 - 6500 |
| L | Absent | 0 - 8 | 1/4 - 2 | Brown | Mount Rainier | 5500 - 6500 |
| N & A | Mount Rainier | 5500 - 6500 | ||||
| O | 1 - 3 | 1 - 3 | Flourlike to fine sand | Yellowish orange | Mt. Mazama | 6800 |
| R | Absent | 0 - 5 | 1/8- 1 | Reddish brown | Mount Rainier | >8750 |
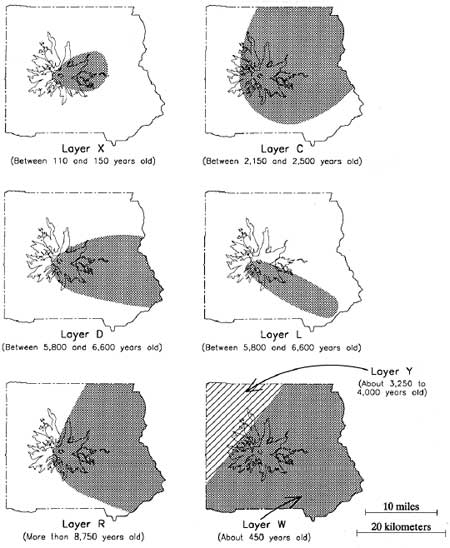
|
| Figure 2.3 Distribution of Selected Volcanic Deposits on Mount Rainier (after Crandell 1987:14) |
Critical points to be made about Mount Rainier's Holocene geology are: 1) the mountain has been extraordinarily active during the time-frame in which humans could reasonably be expected to have used the Park; 2) these events to varying degrees modified the physical structure of the mountain; and 3) these events affected the archaeological record and, to a lesser extent, the character of prehistoric land-use practices. The most dramatic impacts involved destruction of very high elevation landscapes near the summit; and lahar destruction, in filling and subsequent down cutting of major river valleys. Because of extreme elevation, the effect on human use by the former should have been slight.
The impact on river valley land-use practices is more difficult to estimate. Repeated stream flow alteration undoubtedly had a negative effect on native and anadromous fish. Even so, it is likely that fish populations would have returned quickly once affected rivers regained stable streambeds. The primary impact may have been more to resource predictability than to absolute presence/absence per se. The latter is controlled by waterfalls and other major upstream impediments which limited anadromous fish to relatively few Park watercourses. Because of downstream obstructions and lahars emanating from above, human use of Mount Rainier's river valleys is likely to have never been important; particularly when compared to relatively resource-rich subalpine and alpine habitats. Unfortunately, repeated inundation of the floodplains makes riverine use difficult to examine empirically.
None of Mount Rainier's Holocene volcanic deposits were deep enough to have disturbed plant and animal associations in a massive way. Floral and faunal perturbations are likely to have been relatively short lived with insufficient impact on land-use patterns to show in the archaeological record. Even so, the tephra deposits make cultural remains substantially more difficult to locate. During the current reconnaissance, Mt. Mazama tephra was observed between 80 and 100 cm deep in several locations in the Park's NE quadrant. On the south flank of Sunrise Ridge, lithic debris were sandwiched between St. Helens Yn and Rainier C deposits 25 cm below the modern ground surface. While depth of cultural deposits vary with age and position on the mountain, it is clear that efforts must be made to locate prehistoric materials in subsurface context. It is important to recognize that materials documented by surface reconnaissance alone will tend to under-represent actual prehistoric site count, under-represent materials variability and abundance at identified sites, and bias results toward more recent, shallow deposits.
ENVIRONMENTAL ZONES, RESOURCES AND HUMAN
LAND-USE PATTERNS
Ecological Maturity, Resource Abundance and the Northwest Maritime Forest
Due to similar latitude and position relative to Pacific westerlies, the Cascades share general climate and environmental patterns. The Coast Range, Puget Trough, Olympics and Cascades are exposed to a relatively mild, seasonally wet maritime climate along the foothills and western slopes. As Pacific air masses flow over the high Cascades and down the eastern slope, adiabatic warming increases moisture holding capacity and induces the drier climatic regimes characteristic of the Columbia Plateau.
In direct response to moist, relatively stable climatic conditions, the greater northwest maritime region tends toward states of high ecological maturity; that is, toward generally dense, stable forest cover. While not alluding to maturity directly, Heusser (1983:241), citing Waring and Franklin (1979), emphasizes this effect for the Pacific Northwest.
The climatic regimen of winter storms (with protracted periods of moisture and cloudiness, summers with relatively short intervals of soil-water loss, and moderate temperatures over much of the year) is very favorable for tree growth and accounts for the dense coniferous forests of the region. The forests with massive trees of great longevity are unrivaled among temperate forest regions of the world.
In a natural state, dense hemlock and fir forests carpet much the region from the Puget Trough, to the eastern slopes of the southern Washington Cascades. Though specific plant communities vary in response to micro-climate and sediment differences, and especially to forest perturbation (lahars, winds and fires), the general tendency is toward mature, wet forest conditions up to about 5,500 ft above sea level near Mount Rainier. I refer to this mature forest pattern as the Northwest Maritime Forest (see Burtchard and Keeler 1991:17-32). At higher elevations, Pacific Maritime Forests give way to subalpine parkland, alpine tundra, and, on the highest peaks, to glacial snowpack and rubble. On the eastern slope, maritime fir forests grade to more xeric interior pine stands and ultimately to the shrub steppe of the Columbia Plateau.
Forest maturity patterns are important because they are linked to the distribution and abundance of edible plant and animal resources in the maritime Northwest. In a mature or late successional state, Northwest Maritime Forests are difficult places to live. Food sources are limited by the tendency of uniform, high maturity forests to lock up most of their energy and nutrient budget in standing biomass (principally cellulose). A relatively small percentage goes to edible foliage or reproductive parts (see Pianka 1974:89-90; Odum 1971:385; and Whittacker 1975:137). [6] Edible biomass is highest not in the forest proper, but in lower maturity subalpine parkland and alpine tundra associations above the forest line, or at other places where forest cover has been reduced temporarily by fire, landslides, lahars or other sources of disturbance. Consequently, in the Cascades and on Mount Rainier, the most productive habitats over the long term are alpine grasslands, patchy subalpine forest, open wet meadow associations, and patchy interior pine forests (not represented in the Park). Disturbance related low maturity areas are productive as well, but tend to be temporary; closing as the maritime forest regenerates. Laced through this general pattern are salmon bearing rivers and tributary streams locally capable of sustaining intensive human use given sufficient need and labor to meet extraction and storage demands.
In short, high maturity tendencies of the Pacific Maritime Forest tend to suppress food resources necessary to sustain prehistoric populations. In order to maximize access to seasonally available game and edible plants, successful groups were obliged to focus their activities toward lower maturity or earlier succession zones regardless of cultural affiliation or idiosyncratic preferences (i.e., toward open to patchy habitats however they were created and maintained). Accordingly, the key to understanding patterned prehistoric use of Mount Rainier, lies in large measure in our ability to understand the distribution of low to higher maturity habitats within the general forest regime. The following sections address this issue.
Mount Rainier Environmental Zones and Human
Use
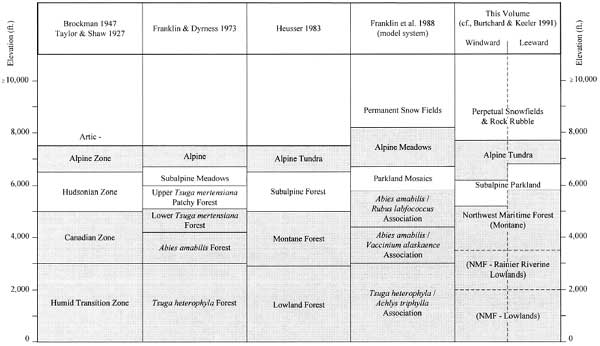
Several taxonomic systems have been devised to characterize forest or plant associations in the Pacific Northwest. The earliest widely applied system characterized forest constituents in terms of south to north latitudinal patterns for the North American continent; keyed to roughly comparable elevation zones on Mount Rainier. In their manuals on Mount Rainier flora and fauna, Brockman (1947) and Taylor and Shaw (1927) used this system to distinguish Humid Transition and Canadian, Hudsonian subalpine forest, and Arctic-Alpine snowfield and tundra zones for the Park. Even though the system oversimplifies local variability, it nonetheless provided useful distinctions between modal plant and animal communities that result from elevationally induced variation in growing season and snowpack.
Perhaps the best known environmental taxonomy for Oregon and Washington was developed by Franklin and Dyrness (1973). Because of its descriptive rigor and applicability to varied settings, this work has become the de facto standard for archaeologically-related environmental descriptions in the Northwest. Franklin and Dyrness' system improves on earlier continental zones by emphasizing combined effects of physiography, climate, and sediment structure to map climax plant associations. For the southern Washington Cascades and Mount Rainier, climax forest zones include 1) Tsuga heterophyla (western hemlock) forest at lowest elevations; grading to 2) Abies amabilis (Pacific silver fir) associations on intermediate slopes; and 3) Tsuga mertensiana (mountain hemlock) forests above 1,250 m (4,100 ft). Mountain hemlock associations become increasingly patchy toward timberline until dominated more by subalpine meadow grasses, sedges and composite flowers than by trees. Fully alpine tundra grasslands are situated between timberline and permanent snowfields above circa 8,000 ft. As will be seen, the system used in the present report builds on Franklin and Dyrness' scheme by grouping Tsuga heterophyla through lower Tsuga mertensiana forests into the single Northwest Maritime Forest class that emphasizes their shared tendency toward high maturity.
In Vegetational History of the Northwestern United States, Heusser (1983:243) models forest associations from the Puget Trough to the crest of the southern Washington Cascades near Mount Rainier. He emulates Franklin and Dyrness' distinctions but couches them in terms more easily recognized as forest/landscape types. Heusser's model (Figure 2.4) is included below because of its similarity to environmental distinctions used here, and because it simulates a mountain profile, drawing attention to the effects of decreasing temperature on key vegetation constituents. Modifications are indicated in brackets.
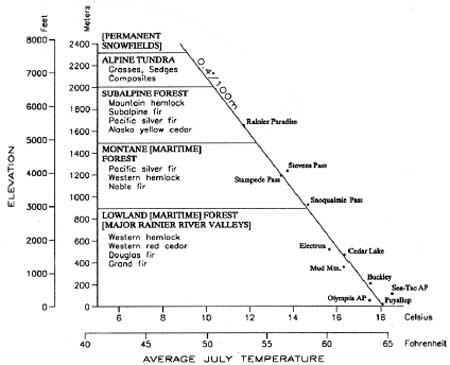
|
| Figure 2.4 Vegetation Zones: Puget Trough - S. Washington Cascades (after Heusser 1983) |
The most thorough consideration of forest association patterns for the immediate project area is Franklin et al.'s (1988) The Forest Communities of Mount Rainier National Park. The authors identify a mosaic of climax associations keyed to four Park quadrants and associated river systems. Elevation and moisture related variability in floral composition and understory dominants are used to provide a more realistic picture of plant associations in various parts of the Park. Importantly, the authors move beyond the three-part high maturity/climax plant association system to consider maturity reducing impacts of major fires, avalanches and lahars. Figure 2.5 is a simplified mosaic of Mount Rainier National Park habitat types as affected by temperature (i.e., elevation) and precipitation (primarily windward versus leeward position on the mountain). It provides a notion of the complexity that underlies simpler forest constructions; and draws attention to an important food resource –Vaccinium – as an understory constituent and seral dominant when Abies amabilis forests are disturbed by fire (see the ABAM/VAAL association in Figure 2.5). Interested readers should refer the complete volume (see also Moir 1989) for detailed consideration of Mount Rainier floral associations.
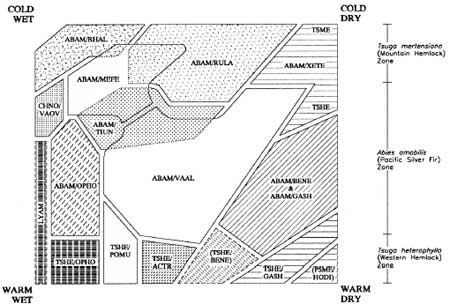
| ||||
Figure 2.5 Generalized Distribution of Mount Rainier Habitat
Types (after Franklin et al. 1988:34)
| ||||
Useful as it is for developing a thorough understanding of Mount Rainier forest ecology, the Franklin et al. (1988) study is unnecessarily complex for present purposes. The most salient issues are those that address relative forest maturity, distribution and abundance of food resources, and factors that alter the forest's maturity/resource balance. It already has been emphasized that in general 1) Northwest Maritime Forests tend toward high maturity; 2) variety and abundance of game and edible plant resources tend to vary inversely with maturity; 3) maturity decreases in a more or less regular, predictable fashion with elevation, maximizing food resource abundance in subalpine and lower alpine habitats; and 4) maturity is reduced in an irregular, less predictable fashion in response to large fires and other forest disturbing phenomena. To accommodate these considerations, the system outlined below isolates five environmental zones that includes three major plant associations keyed to variable maturity and resource distribution patterns; a low elevation river valley subzone; and high elevation glacier/ permanent snowfield category. The maturity reducing effect of fire on forest habitats is discussed as well.
Table 2.2 correlates the present taxonomy with those outlined above. Franklin et al.'s (1988:64-75) modal forest ecosystem structure is used to represent their more elaborate mosaic system. Please note that elevation breaks between all zones are approximate. Boundaries typically are more gradual than shown and vary locally with precipitation, exposure, sediment structure and so on. Alternating gray and white bands indicates zonal concordance to the system used here.
Table 2.2 Mount Rainier Environmental Zonation Systems
Northwest Maritime Forest
Low to Mid-Elevation Forest
Except for high elevation landscapes, Northwest Maritime Forest
associations blanket most of the region from the Puget Trough to the
eastern flank of the southern Washington Cascades. Western hemlock
(Tsuga heterophylla) and western red cedar (Thuja plicata)
are consistent low elevation dominants. Though probably a seral
constituent, Douglas fir (Pseudotsuga menziesii) is presently the
most common tree associated with low to moderate elevation forests.
Salal (Gaultheria shallon) is a frequent understory species
(Franklin and Dyrness 1973:89). Prairies with patchy stands of Douglas
fir, oak (Quercus garryana), and shrubs are common in the Puget
Trough south of Puget Sound. Prior to modern development, these lower
maturity prairies probably were maintained by low summer precipitation
coupled with gravelly glacial outwash soils, and by periodic burning by
Indian groups (Franklin and Dyrness 1973:89; cf., Taylor and Shaw
1927:9). The latter case illustrates the capability of humans to
maintain and/or increase the productive capacity of their environment by
repressing forest succession (i.e., by lowering ecosystem maturity).
Low elevation western hemlock forests grade into mid-elevation associations dominated by Pacific silver fir (Abies amabilis) at about 3,000 to 3,500 ft in the southern Washington Cascades. Silver fir associations are the Park's primary forest type up to circa 4,000 to 4,500 ft. This is the heart of Franklin et al.'s (1988) silver fir/Alaska huckleberry (ABAM/VAAL) and other Abies amabilis associations shown on Figure 2.5. Though dominated by silver fir, forest constituents vary widely with exposure and age. Other common tree species include western hemlock, noble fir (abies procera), Douglas fir, western red cedar (Thuja plicata–wet sides), and western white pine (Pinus monticola). Aside from Vaccinium, locally dominant understory constituents include Oregon grape (Berberis nervosa) and salal (Gaultheria shallon) on the dry side; and devilsclub (Oplopanax horridum) and skunk cabbage (Lysichitum americanum) in wetter areas.
Upper Elevation Forest
Above 4,500 ft, mountain hemlock (Tsuga mertensiana),
subalpine fir (Abies lasiocarpa), Alaska yellow cedar
(Chamaecyparis nootkatensis), and western white pine (Pinus
monticola) silver fir forests become increasingly prevalent. On
wetter northwest, west and southeastern exposures, Pacific silver fir
tends to maintain its upper story dominance. Primary understory
co-dominants, however, shift from Alaska huckleberry toward dwarf
blackberry or dwarf bramble (Rubus lasiococcus), rustyleaf or
fool's huckleberry (Menziesia ferruginea) and rhododendron
(Rhododendron albiflorum) (Franklin et al. 1988:18). On the drier
northeastern, lee side of the mountain, mountain hemlock tends toward
upper story climax dominance, with beargrass (Xerophylum tenax)
dominating low ground cover. Due principally to vegetation repressing
effects of snowpack, closed maritime forests tend to reach only to about
5,200 ft on the wetter windward side of Mount Rainier, and to about
5,800 ft on the drier leeward side.
Forest Fauna
Taylor and Shaw (1927:11) list animals common to Mount Rainier's low
to mid elevation forests as
...Harris woodpecker, Gairdner woodpecker, northern red-breasted sapsucker, western pileated woodpecker, western flycatcher, Steller jay, Louisiana tanager, chestnut-backed chickadee, western golden-crowned kinglet, russet-backed thrush, and western bluebird. Mammals ...are the Trowbridge shrew, brown bat, bobcat, Cooper chipmunk, beaver, and snowshoe rabbit.
Of the larger animals, black-tailed deer are present in the forest were adequate browse is available, particularly at meadow fringes. However, both deer and elk (presumed to be present prehistorically) "during the summer months ...seek the higher levels of the park, where open spaces afford more sunlight and grass. They [deer] are especially fond of burns, where the thick vegetation provides an abundance of browse" (Taylor and Shaw 1927:118). Similar considerations apply to black bear. Marmots, goats and large birds (grouse and especially ptarmigan) tend to frequent higher elevation subalpine to alpine landscapes.
It is interesting and, from a prehistoric hunter's point of view, important to note that the most abundant mid-forest animal species are relatively small birds and mammals. Even though more economically useful larger game is present where browse permits, over the long run, most successful hunters would have followed their prey through the forest into more open subalpine and alpine settings at higher elevation.
The Huckleberry Issue
The single most noteworthy exception to the food resource poor
qualities of mature mid-elevation montane forests lies in their response
to major maturity-reducing disturbances. Franklin et al. (1988:150) note
that "Wildfire is, by far, the most important agent of forest
catastrophe at Mount Rainier National Park, followed by snow avalanches
and debris flows (lahars) (Hemstrom and Franklin 1982). Wildfires have
occurred on approximately 90 percent of the existing stands, avalanches
on 7 percent, and lahars on 2 percent." In cases of forest catastrophe,
huckleberries are a common early successional dominant. According to
Franklin et al. (1988) Vaccinium alaskaense is usually the most
abundant of the Mount Rainier huckleberries. [7] Four other mid-elevation varieties also are
common–black or big-leaf (V. membranaceum), blue or
oval-leaf (V. ovalifolium) and red (V. parvifolium)
huckleberry, and the small red grouseberry (V. scoparium).
Cascades or dwarf blue huckleberry (V. deliciosum) is more common
higher in subalpine settings. Western bog blueberry (V.
occidentale) and evergreen huckleberry (V. ovatum) are found
in low abundance, at low elevation (see Brockman 147:127-128). Abundance
of all species is increased by maturity reducing forest disturbances,
particularly fire, but diminishes as forest cover returns.
Because of its edible resource qualities and because deer, bears and birds are attracted to productive huckleberry grounds, Vaccinium may have been important to prehistoric foraging strategies. Indeed, during early historic times, huckleberry collection appears to have been the primary reason Indian groups came to Mount Rainier (Smith 1964:155). The most important varieties reported by Smith (1964:157-163) were oval-leaf (probably including Alaska) and big-leaf huckleberries in midelevation forests; and low growing dwarf huckleberries at higher elevation. Among other things, Smith goes on to discuss huckleberry drying techniques on mats and elevated racks; popular collecting grounds; the practice of setting fires to enhance huckleberry production; and packaging for downslope transport. At several points he notes the use of horse transport and coordination of berry collecting with hunting activities.
In my opinion, it is doubtful that the nutrient or caloric value of huckleberries was sufficient to overcome distance and bulk transport difficulties associated with mass harvest prior to introduction of the horse circa A.D. 1700. There is no doubt, however, that the resource was used extensively by small collecting parties emanating from various tribal groups on all sides of the mountain thereafter (see Smith 1964:149-179). Furthermore, given its history of repeated burns prior to modern fire suppression (Franklin et al. 1988:149-158), Mount Rainier could have supported substantial huckleberry habitat at ostensibly resource poor maritime forest elevations. For now, it must be recognized that burned and excessively drained mid-elevation habitats may have produced floral and faunal resources sufficient to attract significant human use in the past. The extent to which such use extended into the prehistoric past depends on the direct value of these berries as food plus the value of game co-occurring at midelevation berry grounds, compared to resources found at higher-elevation habitats. I suggest that despite presence of mid-elevation huckleberry habitat, the greater stability and overall higher productivity of subalpine habitats would consistently have attracted greater human use over longer stretches of time. If so, most of the Park's prehistoric archaeological record should be located in subalpine and alpine habitats with a lower frequency elsewhere.
Low Elevation Rivers and Floodplains
In a sense, rivers and associated floodplains are not a distinct environmental zone per se, but rather are more characteristic of maritime forest stands near sea level. This is generally true of the Park's lowest major floodplains–particularly the Carbon, Nisqually and Ohanapecosh Rivers in the Park's northwest, southwest and southeast corners. Brockman (1947:3) enthusiastically describes the Park's lower riverine forest in his Humid Transition Zone introduction as
...awe-inspiring in its quiet solitude; cathedral-like in its sombre, peaceful grandeur. So dense are the trees that their branches interlace overhead to form an evergreen canopy, and even on bright summer days a condition of semi-twilight exists on the forest floor. The great trunks rise from a tangled mass of shade-loving plants which, in places, approach tropical-like luxuriance. Other trees, leveled by age or the elements, sprawl upon the ground–some newly fallen, others festooned with moss, and some in the last stages of decomposition. The trails, carpeted by an accumulation of humus and forest litter, are soft and yielding to the tread.
Lahars and Floods
Inviting as such scenes may be, they are limited in scale by the
Park's overall high relief and physiographic instability. The subzone is
distinguished here less for its unique floral qualities in the mature
state described by Brockman, and more in recognition of Mount Rainier
river valleys' frequent exposure to maturity-reducing lahars and high
energy runoff, and because of presence and potential resource importance
of anadromous fish in some of the river systems.
Lahars represent the most spectacular destructive events to impact Mount Rainier's low elevation riverine environments. Effects of major Mount Rainier mud and debris flows such as the 5,000 year old Osceola and 500 year old Electron mud flows are well known. Lesser flows are destructive on a smaller scale, and are far more common. Because of its relief, heavy rainfall and geological youth, many of Mount Rainier's rivers and streams are subject to periodic disruption, particularly in upper to mid-level drainages (see Crandell and Mullineaux 1967, and Scott et al. 1995). Such events repress forest maturity, temporarily opening affected floodplains to more productive early succession plant and animal associations. Even in the absence of such spectacular events, high energy seasonal runoff on major rivers–such as that shown in Figure 2.6 on the Carbon River–are capable of altering river courses and transporting boulder-sized rock substantial distances, and in so doing, keep the forest fringe at bay.

|
| Figure 2.6 Carbon River Floodplain. (Facing upstream from Northern Loop Trail crossing, September 4, 1995. Note that high energy flow is infringing on the west-bank forest margin.) |
Anadromous Fish
Because they improve forage by suppressing floodplain forest cover,
lahars and high energy floods promote a modest gain in exploitable
resource abundance. The most important riverine resources, however,
would not have been ungulates but salmon. Because of predictability and
high seasonal abundance, salmon has been an important resource for
regional populations since the mid-Holocene. The abundance and extent to
which anadromous fish penetrated into Mount Rainier National Park,
however, remains unclear. Despite repeated floods and debris flows, at
least limited salmon populations have been reported historically in
major river systems within Park boundaries (Samora pers. com. 1995). Her
sources allude to coho salmon (Oncorhynchus kisutch) in the
White, Carbon, Puyallup and Mowich Rivers; chinook (Oncorhynchus
tshawytscha) in both White River forks; and steelhead
(Oncorhynchus mykiss) in the Carbon, White, Puyallup and
Nisqually Rivers. Fulton (1968) reports fall chinook in the Cowlitz
River just south of Park Boundaries. He also reports steelhead spawning
areas in the Ohanapecosh well within the Park (Fulton 1970). Bull trout
(Salvelinus confluentus) and/or Dolly Varden (S. malma)
historically also were present in several of the Park's rivers (Samora
pers. com. 1995).
Combining information above with maps showing spawning areas (Fulton 1968, 1970) and major upstream impediments (Williams et al. 1975), it is possible to reconstruct roughly the extent to which spawning salmon physically could have entered Mount Rainier rivers. Figure 2.7 shows river systems and approximate upstream limits of unimpeded salmon migration. Rivers not highlighted have major cascades or falls that effectively preclude upstream migration well below Park boundaries. Please note that this projection should be considered a working model to be improved as more information is made available.
|
| Figure 2.7 Potential Salmon-Bearing River Systems in Mount Rainier National Park |
Randall Schalk (pers. com. 1996) has studied Fulton and Williams map data, and offers these observations:
The White River appears to have no obstacles below a point slightly above the mouth of Deadwood Creek. The highest for present chinook usage appears to be Pinochle Creek and lower Huckleberry Creek north of the Park. Within Park boundaries, there are several White River tributary creeks that have salmon usage listed as unknown. Many say none. Coho usage is possible in some of the creeks that flow into lower Huckleberry Creek (a few miles north of the Park boundary) and possibly Silver Creek.
No modern salmon usage is shown in the Puyallup River within the Park. LeDout and Niessen Creeks show some coho usage but they are well downstream from the Park. There are three upper basin tributaries that penetrate the park apparently without natural obstructions (source Williams et al. 1975):
South Mowich R. (obstacle shown @ 3 mi into park)
North Fork Puyallup (obstacle shown at @ mi into Park)
South Fork Puyallup (blockage shown at @ mi into Park)
I have no data on steelhead usage of the Puyallup and Mowich Rivers although steelhead probably could get wherever chinook and coho go. Spring chinook probably migrated at least as far up as the cohos and probably beyond.
The main stream of the Nisqually presently is blocked above LaGrande by an impassable dam. How far salmon got up this drainage historically is less clear. No natural obstacles are shown on the river until just below Paradise. Goat and Tenas Creeks have no obstacles shown. Tahoma Creek has none on the lower two miles. No major obstacles are shown on Kautz Creek., though glacial silt is likely to have hindered spawning (source Williams et al. 1975).
Fulton's (1970) steelhead map for the upper Cowlitz shows steelhead presently use the lower portion of the Ohanapecosh River. Coho are shown as using the lower parts of Butter Creek and Lake Creek–two tributaries of the Cowlitz that enter below the Ohanapecosh. Fall chinook are shown in the Cowlitz still further below (Fulton 1968). I [Schalk] suspect that spring chinook probably would spawn at least as far upstream as coho and steelhead although Fulton doesn't provide detail at this fine scale.
It is important to recognize that, because of extensive habitat alteration, present fishery data are not good indicators of prehistoric presence or abundance. We have emphasized the position of upstream barriers to attempt to show maximum possible migration routes. Unfortunately, we cannot reconstruct specific upstream migration patterns or seasonality with precision. Even so, it unlikely that salmon were ever a major resource in the Park. Periodic floods and debris flows temporarily, but repeatedly, must have diminished anadromous fish habitat. Perhaps more important is the high energy, silt-laden character of the near mountain river systems. According to Schalk (pers. com. 1996):
The heavy loads of glacial silts further diminish the value of these kinds of streams as spawning habitat. Some salmon may have penetrated the Park in very limited numbers but salmon fishing this far upstream I believe would have always been poor at best. If people were harvesting any salmon at all within the park I suspect it was infrequent and incidental to other activities.
Floodplain Archaeology and Land-Use
Aside from maintaining
modest low maturity stream side ungulate habitat, the primary impacts of
river dynamics and lahars may have been most pronounced on the
archaeological record itself. Destructive impact of larger lahars is
obvious. Though less destructive, smaller mud and debris flows, and
annual hydrological activity also have erased the stream side
archaeological record from time to time. Accordingly, it is unlikely
that low elevation floodplains preserve a substantial record of the more
distant prehistoric past, at least for larger, more geologically active
drainages.
Even so, there is reason to believe that river floors were not a major element of Mount Rainier's prehistoric resource base. The river valley landscapes experience resource limitations common to maritime forests generally. Indeed, because Mount Rainier's river valleys are too low for productive huckleberry habitat (at least for commonly collected varieties), too narrow for extensive ungulate habitat and too silty and lahar prone for productive anadromous fisheries, their usefulness to prehistoric populations may have been more for access to Mount Rainier itself, rather than for resources offered directly.
Subalpine Parkland
Up from the floodplains and above the maritime forest, dense forest cover gives way to clumped tree groups scattered among meadow communities. Sometimes referred to as the forest/tundra ecotone (Franklin and Dyrness 1973:248), these subalpine parklands typically appear as patchy associations of conifers interspersed with grassy meadowlands sandwiched between closed forest below and open tundra above. Because of high resource potential, and because they compromises distance between tundra and forest ecozones, I suggest that Mount Rainier's subalpine parklands are the most important places for focused seasonal human use during the prehistoric past–clearly not the only places, but the most intensively and repeatedly used over the long-term.
The subalpine zone's enhanced productivity lies in the maturity suppressing effect of heavy snowpack and short growing season. Under such circumstances, rapidly growing and reproducing species such as grasses, sedges, composite flowers, low shrubs, and plants with underground bulbs have an advantage over slower growing, longer-lived trees. Rapid growth plants also provide the best forage for ungulates as well as smaller mammals and birds. Locally dense dwarf huckleberries are good bear and bird habitat and can be exploited directly by humans. Subalpine parklands offer an effective compromise between closed forest and open space–meadows provide a good source of ungulate forage and edible flora, and tree patches provide protective shelter from predators and weather.
Talyor and Shaw (1927:12-13) also emphasize the subalpine zone's unique floral and faunal abundance.
...the Hudsonian Zone (above the narrow forested area), with its happy combination of open grassy park and subalpine flower garden on the one hand and pinnacle and glacier on the other, is without doubt the most attractive zone on the mountain. The shade of the deep Canadian forest is gone. Groups of handsome spirelike alpine firs and mountain hemlocks are set about the meadow borders or interspersed over the green expanses, always affording plenty of light and moisture to the grass and flowers. There is an abundance of food for birds and mammals, which are more conspicuous and probably more numerous in this zone than in any other in the park. The Hudsonian is the most desirable and the only favorable zone for camping away from the hotels, because only here (except at Longmire) can forage for horses be found [emphases added].
Closed Pacific maritime forests give way to patchy subalpine associations above about 5,200 ft on the wetter northwest, west and southern sides of the mountain; and above 5,800 ft on the drier northeastern side. Tree groups become increasingly patchy and stunted with elevation, ultimately giving way to scrubline vegetation and full alpine tundra at about 6,200 and 6,800 ft on windward and leeward sides respectively. Figure 2.8 shows subalpine associations at circa 5,400 ft in Mist Park in Mount Rainier's northwest quadrant. The view is taken from Mt. Pleasant Rockshelter site FS 72-02 (45PI433). Two additional prehistoric sites and two isolate finds have been documented in the near vicinity.

|
| Mist Park Subalpine Meadows from Mt. Pleasant Rockshelter. East-northeast facing photo shows Sluiskin Mountain (jagged peaks at upper left-center), and Old Desolate Mountain, in the background. |
Patchy forest associations are dominated by Pacific silver fir on wetter sides of the mountain. Mountain hemlock tends to predominate on the dry side (see Figure 2.5). Typically, subalpine stands incorporate a higher frequency of mountain hemlock, subalpine fir (Abies lasiocarpa), and white bark pine (Pinus albicaulis) than present at lower elevation. Alaska yellow-cedar is locally present in low density. Near timberline, subalpine fir is one of the last tree species to give way to alpine tundra.
Subalpine Meadows
While tree patches are important for
shelter, our primary interest in subalpine and tundra habitats lies in
the productivity of their meadow communities. As with forests,
composition of subalpine meadows varies with sediment structure,
drainage, elevation, and solar exposure. Citing Henderson (1973),
Franklin and Dyrness (1973:252) identify five major meadow groups on
Mount Rainier: 1) Phyllodoce empetriformis-Vaccinium deliciosum
(red heather-dwarf huckleberry); 2) Valeriana sitchensis (lush
herbaceous); 3) Carex nigrians (dwarf sedge); 4) rawmark and low
herbaceous; and 5) Festuca viridula (grass or dry grass) (cf.,
Whitney 1983:42-43). Community associations within these groups is
highly variable, though here we focus only on dominant constituents.
Heather-dwarf huckleberry associations (Group 1 above) are most prevalent on gentle slopes with moderately well-drained soils. A good example are the south trending ridge near Mt. Ararat and parts of "Indian Henry's Hunting Ground"in the Park's southwest quadrant. Both heather and huckleberry are low-growing shrubs capable of forming nearly complete low ground cover mats. Franklin and Dyrness (1973:256) suggest that more purely dwarf huckleberry stands (such as those near Mt. Ararat) tend to be pioneer communities on burned habitats, and are succeeded either directly by forest (at lower elevation) or by more complex heather-huckleberry associations.
Ethnohistoric reports on the importance of dwarf huckleberry (V. deliciosum) are contradictory. Smith's (1964:160-161) sources refer to it variously as 1) of little economic importance because it grows too low to the ground and ripens too late in the year; and 2) gathered in quantity because the berry is large and low to the ground. It is plausible that the apparent contradiction is due to use as a buffering resource in years when lower elevation, taller V. alaskaense, ovalifolium and membranaceum varieties were not productive. In any case, dwarf huckleberry's subalpine co-occurrence with hunted game is likely to have ensured use minimally as a supplemental, if not staple, food item.
The lush herbaceous meadows (Group 2) are the richest and most diverse of the subalpine meadow communities (see Franklin and Dyrness 1973:256-262). These meadows develop on steep to moderate gradient, well watered, avalanche-prone slopes. They are common to the southern and western slopes of Mount Rainier. Specific floral composition varies. Community dominants listed by Henderson (1973) in Franklin and Dyrness include varieties of valerian (especially Velariana sitchensis), false hellebore (Veratrum viride), lupine (Lupinus latifolius), bistort (Polygonum bistortoides), sedge (Carex spectablilis), and monkeyflower (Mimulus lewisi). Actual composition, of course, is substantially more diverse. More important than specific floral composition is the capacity of these low maturity associations to provide forage for a variety of exploitable mammals such as elk, deer and marmots. If continuously maintained by frequent landslides, such communities may have been of substantial importance to prehistoric hunter-gatherers.
Black sedge (Carex nigricans) communities (Group 3) typically are found on relatively flat to rolling sites with short growing season due to late persisting snowpack and cold, wet soils (Franklin and Dyrness 1973:261). Floral diversity is relatively low and growth limited by the circumstances noted above. Local co-dominants include marshmarigold (Caltha biflora) and pedicularis (Pedicularis groenlandica).
Franklin and Dyrness' rawmark and low herbaceous category (Group 4) consists of poorly developed, pioneer associations in a variety of locations with well drained bouldery/gravely to sandy sediments (Franklin and Dyrness 1973:262-263). Slope is variable, ranging from alpine scree to low gradient regisols. Total plant cover varies from a low of about 20 percent to nearly complete. Dominant associations include saxifrage (Saxifraga tolmiei), luetkea (Luetkea pectinata), buckwheat-pussypaws (Eriononum pyrolaefolium-Spraguea umbellata), everlasting (Antennaria lanata), and aster-everlasting (Aster alpigenus-A. lanata).
Grassy parklands (Group 5) are particularly well developed on the drier northeastern side of the mountain. Two dominant community types intergrade on a moisture gradient (Henderson 1973 in Franklin and Dyrness 1973:263): fescue-lupine (Festuca viridula-Lupinus latifolius) in moist locations and fescue (F. Viridula-Aster ledophyllus) in drier places. Fescue-lupine associations are most commonly associated with level to low gradient parklands such as Sunrise Ridge and Grand Park. Fescue-aster associations are found on even drier steep, south facing slopes with coarse textures soils.
Subalpine Fauna
Subalpine communities were important
prehistorically both because they produced directly consumable plants
(e.g., huckleberries and alpine lily), and because they supported
relatively high bird and mammal populations as well. A sample of common
species can be gleaned from Taylor and Shaw (1927:13), Whitney
(1983:42-43) and Schamberger (n.d.:21-23) summary accounts. The birds
reported by these sources include golden eagle, saw-whet owl, calliope
hummingbird, Clark nutcracker, Cassin purple finch, western sparrow,
warblers, mountain chickadee, Townsend solitaire, hermit thrush,
mountain bluebird, northern three-toed woodpecker, Hammond and
olivesidedflycatcher, golden-crowned kinglet, gray jay, red-breasted
nuthatch and evening grossbeak. Mammals include red fox, Washington
weasel, Canada lynx, mountain lemming mouse, Rainier meadow mouse,
large-footed mouse, red-back, Oregon and heather voles, Rainier pocket
gopher, hoary marmot, yellow pine and Townsend chipmunk, mantled ground
squirrel, snowshoe hare cony (pika), pine marten, coyote, black bear,
mountain lion, elk and black-tailed deer. Though more common to alpine
tundra, mountain goats occasionally graze in upper elevation subalpine
associations and move through them to upper forest zones to find winter
shelter. Ptarmigan, common in tundra associations, also are found in
subalpine places.
At first glance, faunal variation may not appear markedly different from the forest proper. However, overall diversity is greater and, importantly, seasonal abundance of larger and fatter (i.e., energy rich) animals is substantially higher. Animals such as black-tailed deer, elk, black bear, and marmots find better forage in the meadow/forest ecotone than in the closed forest (except for burn areas). Assuming game animals are not over hunted, the subalpine combination of high return faunal and floral species should have been attractive to human exploitation throughout the prehistoric past.
Subalpine Parkland, Land-Use and Archaeology
The primary
point to be taken in the discussion above is that subalpine communities
substantially increase the abundance and diversity of low
maturity flora over that of lower elevation maritime forests. Enhanced
forage, in turn, favors increased faunal abundance. Assuming humans tend
to structure their annual round to seek out productive habitats, we
should expect subalpine parklands to have been the primary focus of
human hunting and gathering activity throughout the prehistoric past.
While burns and landslides also tend to be resource rich, only subalpine
and alpine zones do so in a manner that is stable over a long period of
time. If subsistence resource availability was the principal variable
attracting prehistoric people to Mount Rainier, it is reasonable to
expect Mount Rainier's archaeological record to reflect a bias toward
low maturity subalpine to alpine landscapes.
It should be noted in closing that subalpine communities exist in a state of dynamic tension between lower forest boundaries and the upper tree or shrubline (krummholtz) boundary with alpine tundra (see Franklin and Dyrness 1973:276-284). In the subalpine ecotone, density of tree stands decreases with elevation as dictated by local effects of wind, fire, avalanches and snowpack; and by the overarching effect of decreasing temperature. In general, trees invade subalpine meadows during prolonged periods of stable warm weather, and retreat in the face of cold or unstable conditions. That is, parkland meadows tend to shrink and move upslope during periods of climatic warming (providing fire frequency does not increase); and expand and move downslope during cool, excessively dry and/or unstable circumstances. If subalpine communities are as important to prehistoric hunters and gathers as suggested here, it is important to develop a thorough understanding of Holocene climatic variables in regard to their implications for the resource productivity of these zones.
Alpine Tundra
Upper elevation tree clumps and scrubline mats of Englemann spruce (Picea englemannii) and subalpine fir give way to open alpine communities at about 6,200 to 6,800 ft on Mount Rainier. Alpine tundra extends upward to the firnline boundary with permanent snowfields at circa 7,600 ft. Because of Mount Rainier's great height and geological youth, much of the circa 7,600 ft upper boundary with permanent snowfields is characterized by rock and essentially vegetation free glacial rubble.
Figure 2.9 shows alpine tundra habitat and a pair of mountain goats (Oreamnos americanus) on Burroughs Mountain in the northeastern side of Mount Rainier. Fryingpan Glacier in the background and lower reaches of Emmons Glacier at photo right show increasingly barren glacial scree slopes and glacial scour at the alpine interface with standing snowfields.

|
| Figure 2.9 Mountain Goats on Burroughs Mountain Alpine Tundra. (South facing photo shows Fryingpan Glacier and White River headwater snow fields in the background.) |
Alpine Tundra Flora
The greatest expression of alpine communities on Mount Rainier is on the
drier northeastern slope at places like Goat Island Mountain, Burroughs
Mountain, and terrain near Frozen Lake and Mt. Fremont. The growing
season is short with substantial diurnal temperature variation. Terrain
is open with tundra grasses and forbes growing at moderate density over
variously exposed volcanic lapili and shallow lithosols. Specific plant
associations vary with local conditions. At least at lower elevations,
subalpine meadows extend with minimal constituent variation into the
open tundra. Dominant plant associations on the northern slope were
studied by Hamann (1972) and reported by Franklin and Dyrness (1973:287)
as follows:
Black crowberry/prairie lupine (Empetrum nigrum/Lupinus lepidus); north-facing fellfields.
Kinnikinnik/coast goldenrod (Arctostaphylos uva-ursi/Solidago spathulata); south-facing fellfields.
Blunt-leafed sandwort/prairie lupine (Arenaria obtusiloba/L. lepidus); pioneer on gentle slopes.
White heather/alpine aster (Phyllodoce glanduliflora/Aster alpigenus); moistest alpine environments.
Coiled pedicularis/showy sedge (Pedicularis contorta/Carex spectabilis); forms well-developed turf.
Golden fleabane/prairie lupine (Erigeron aureus/L. lepidus); forms well-developed turf.
Spreading phlox/mountain sandwort (Phlox diffusa/Arenaria capillaris); seral to pedicularis/sedge.
Alpine Fauna, Land-Use and Archaeology
Floral and faunal
diversity decreases above the scrubline. Successful animal species are
those suited to grazing on the low ground flora, cope with unstable
weather without tree shelter, and make use of high elevation snowfields
and cleavers for predator escape. Mountain goats, pika, white-tailed
ptarmigan, marmots and other smaller rodents are common alpine tundra
feeders during its brief July through September summer. Ravens, larks,
finches and piptis (see Taylor and Shaw 1927:15; Whitney 1983:43-44;
Schamberger n.d.:22-23) also use subalpine habitats. While extirpated
from the Park by the late 1800s or earlier, elk are also likely to have
been warm season grazers in the lower alpinesubalpine boundary. Because
of the brevity of alpine summers, by early to mid October, game becomes
virtually unavailable. Pika and marmots move to underground dens and elk
migrate to lower-elevation forage. Goats move downslope a short distance
to upper forest wintering areas such as Tum Tum Peak on Mount Rainier's
southwest slope (Schamberger n.d.:75).
It is likely that goats, elk and perhaps marmots provided sufficiently productive food and wool sources to attract human predation in the alpine tundra zone. In the historic period, goat wool was highly prized for its beauty and warmth (see Eells 1985). Indeed, though other animals undoubtedly are present, to date the only unambiguously identified faunal remains in archaeological context are goat teeth from Fryingpan Rockshelter (FS 63-01 [45PI43]) (Gustafson:1983:27-28). Marmots may have been important for their fat. The benefit of elk, of course, lies in large body size and correspondingly high meat return per animal killed.
Because of unstable weather and availability of fewer specific resources, it is probable that alpine foraging tended to be carried out by small task-specific hunting groups with short duration camping, ambush, butchering or lithic procurement stops. If so, we should expect the alpine/upper subalpine archaeological record to reflect such constraints in the form of more functionally limited tool kits and lower artifact density (except at lithic quarries and butchering sites such as FS 86-01 [45PI407] at Frozen Lake). These qualities contrast with the subalpine to subalpine forest boundary where resource variety is greater and weather is milder. Lower-elevation conditions favor longer-term use, probably by a higher frequency of mixed sex gathering and hunting parties. We expect subalpine/upper forest margin sites should exhibit the full rage of sites common to alpine habitats plus a number of higher artifact count, multi-task assemblages consistent with more sustained and varied activities, and probably with more repetitive use. It is plausible, too, that select lower elevation places functioned as base camps from which more limited task gathering and hunting parties operated. Indeed, that argument will be made for three of Mount Rainier's lower subalpine/upper forest sites–FS 95-10 (45PI429) in the Park's southwest quadrant, and FS 71-01 (45PI438) and FS 90-01 in the northeast quadrant.
Perpetual Snowfields and Glaciers
Perpetual snowfields with standing glaciers and associated barren rubble fields range from firnline at 7,600 ft to Mount Rainier's 14,410 ft summit. Individual glacial tongues and young recessional moraines extend downslope in major glacial valleys to circa 5,500 ft. The largest of these–Carbon, Winthrop, Emmons, Ingraham, Nisqually, Tahoma and Mowich Glaciers–extend to below 5,000 ft.
Above the firnline, vegetation is suppressed by snow and ice mass. Floral species are limited essentially to mosses and lichens with very few grasses and flowering plants on rocky cleavers and other seasonally ice-free areas (Brockman 1947:6). Fauna are limited as well. A few marmots, pika, mice and shrews have been observed as high as 8,000 to 10,000 ft. Mice and even bear tracks have been reported at the summit (Schamberger n.d.:22). Clearly, such events are rare, and high mountain edible biomass is negligible. Perhaps the single exception to the pattern the presence of mountain goats which take advantage of glaciers and high rock fields for travel, observation, and escape. For the most part, however, Mount Rainier's glacial zone is largely devoid of economically useful plant and animal life.
Given logistical difficulty, relatively high risk and low return, we assume that prehistoric human use of high elevation Mount Rainier was limited to goat hunting, possible ceremonial use of the alpine/glacier interface, and very infrequent use of higher elevation snow and boulder fields. The highest elevation prehistoric archaeological finds presently documented on Mount Rainier are two isolated projectile points found at 7,500 ft on Success Cleaver (IF 01-70 in the southwest quadrant). Their presence is consistent with use of the locality as a hunting stop at the upper alpine/lower permanent snowfield boundary. We expect similar finds, perhaps associated with low stacked rock features or talus pits to be documented as cultural resource studies continue.
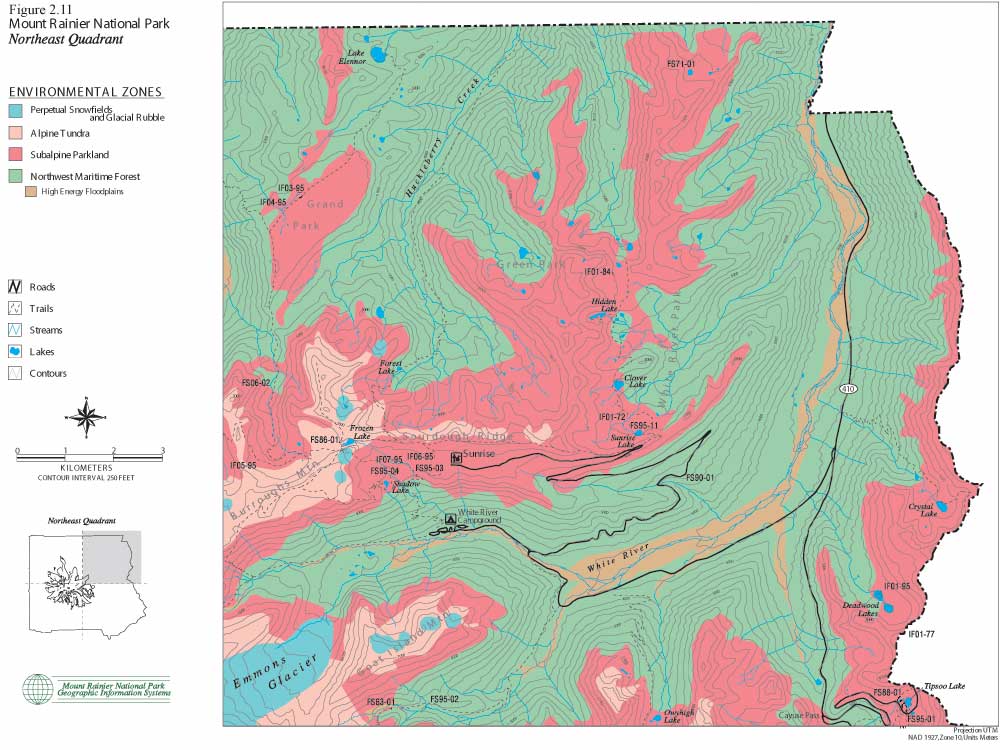
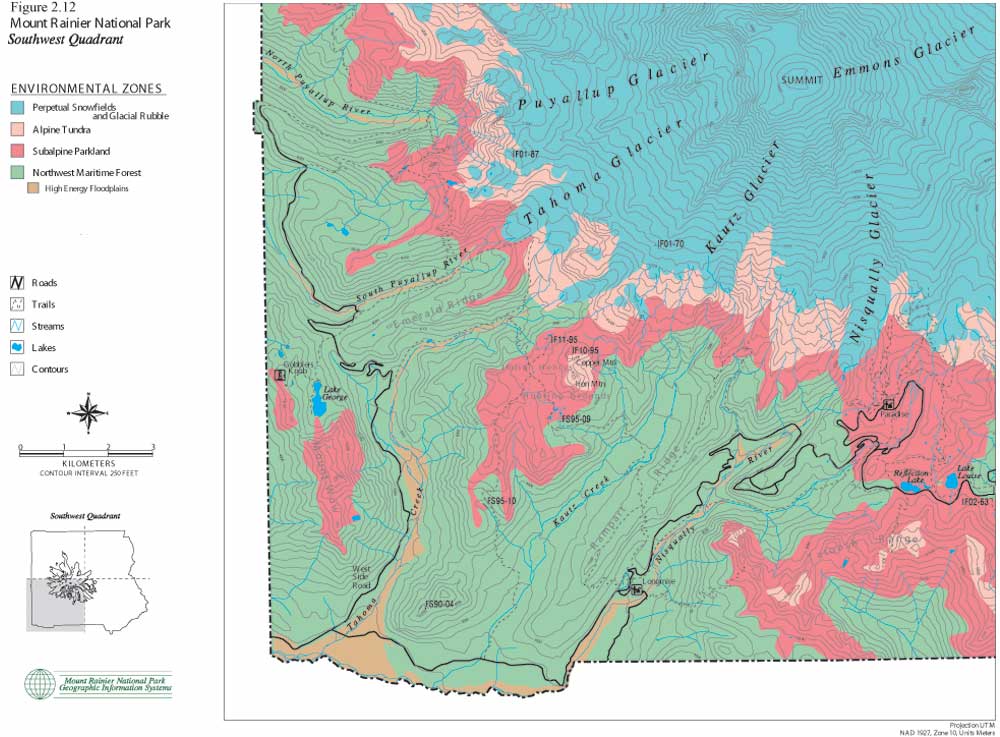
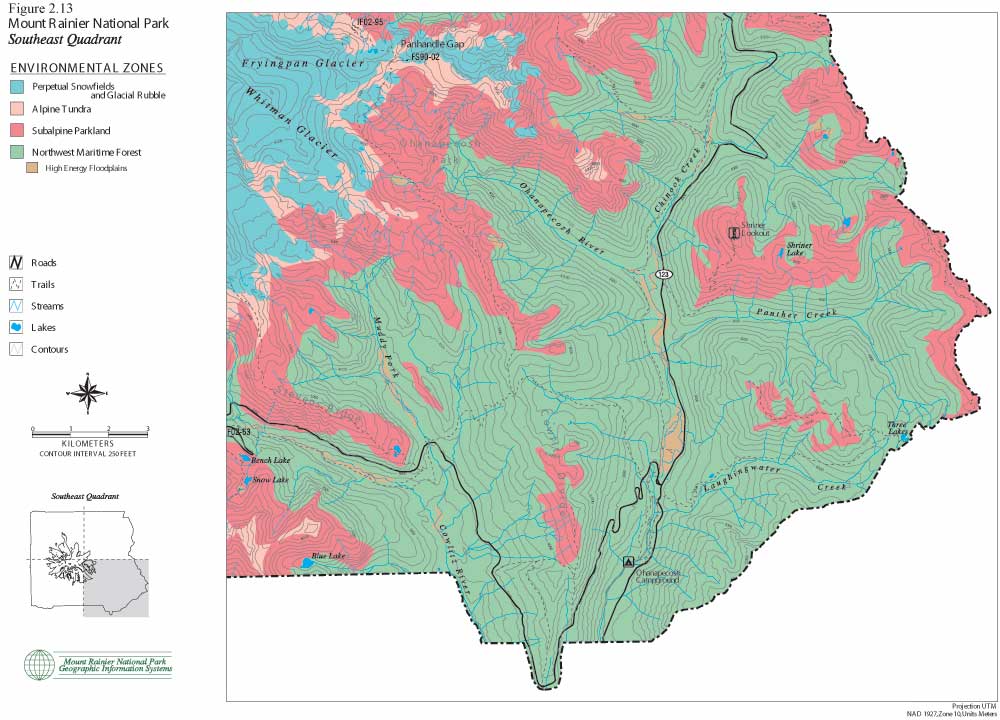
The following set of color maps (Figures 2.10, 11, 12 and 13) show Mount Rainier environmental zones and formally documented archaeological sites in four Park quadrants. Zonal boundaries are derived from environment data logged into the Park's geographic information system (GIS). Basic data on forest, subalpine and alpine boundaries were plotted from aerial photographs. They are intended to model climax vegetation associations, excluding effects of short-term fire alteration. Perpetual snowfield boundaries include standing glaciers, permanent snowpack and a 100 ft buffer to accommodate glacial scree slope rubble. High energy floodplains are those that have experienced major documented floods and/or lahars. These maps provide the best precision presently available for displaying environmental zones and for graphically illustrating association with documented prehistoric sites. The clear association of prehistoric sites with subalpine and alpine settings is consistent with ecological land-use arguments offered above, in ethnohistoric accounts collected by Smith (1964), and discussed in the following sections.
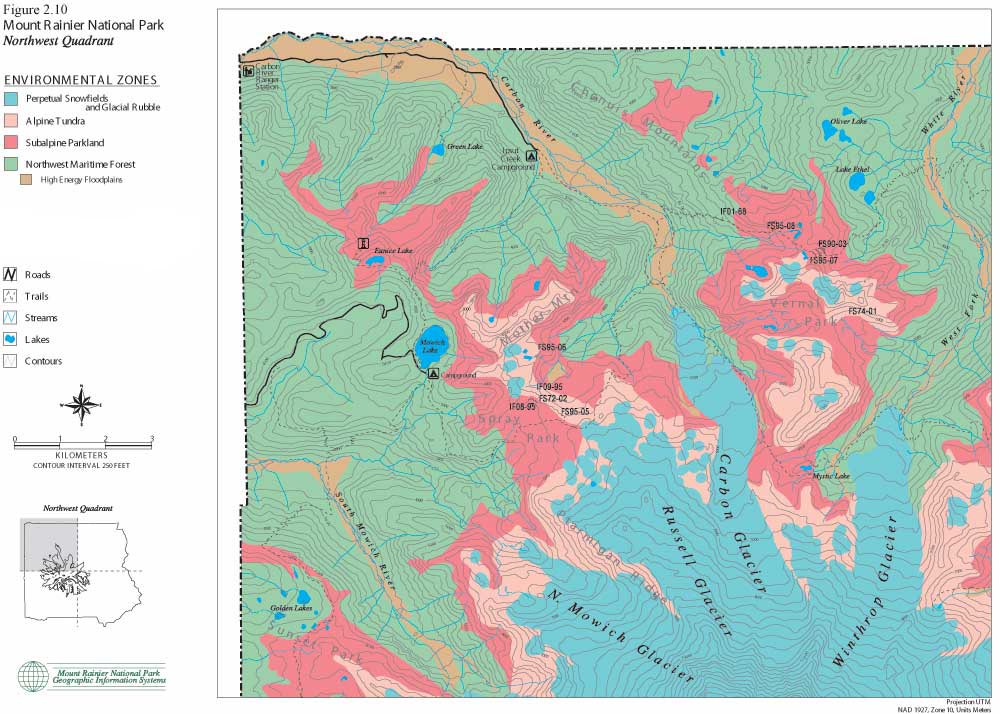
|
| Figure 2.10 Mount Rainier National Park. Northwest Quadrant. (click on image for an enlargement in a new window) |
|
| Figure 2.11 Mount Rainier National Park. Northeast Quadrant. (click on image for an enlargement in a new window) |
|
| Figure 2.12 Mount Rainier National Park. Southwest Quadrant. (click on image for an enlargement in a new window) |
|
| Figure 2.13 Mount Rainier National Park. Southeast Quadrant. (click on image for an enlargement in a new window) |
HOLOCENE EVOLUTION OF MOUNT RAINIER'S ENVIRONMENT
The central premise underlying environmental and land-use patterns discussed above holds that critical plant and animal resources are not evenly distributed but rather tend to cluster seasonally in 1) upper-elevation subalpine to alpine habitats, and 2) early fire succession habitats in mid-elevation forests. Assuming that these plant and animal resources are the primary attractors drawing lowland populations to the mountain, it is reasonable to expect human use to have always favored these places over other less productive forest zones. Because of their spatial stability (relative to burn areas) and because they optimize access to the widest range of exploitable resources, subalpine to alpine habitats are expected to have been particularly heavily used during the prehistoric past.
Vegetation boundaries are not perfectly constant over periods. There is little doubt that Holocene climatic changes altered high elevation forest patterns, affected resource productivity, and may have altered human land-use strategies on Mount Rainier. At a general level, there is ample reason to believe most of the northern hemisphere experienced general climatic warming following final retreat of Fraser/Wisconsin glacial epoch about 9,500 years ago. The trend persisted until about 4,500 to 4,000 years ago, then reversed, with generally cooler conditions to the present.
Even though this pattern substantially simplifies actual Holocene climatic oscillations there is reason to expect climatically induced variation in forest and resource structure during the period of time that humans have inhabited the Mount Rainier region. Without active human intervention, periods of sustained environmental warming could be expected to be accompanied by forest advance in upper elevation environments, effectively reducing the area of economically productive subalpine habitat. Alternatively, delayed snowpack melt associated with sustained colder periods would have repressed montane forest cover, ostensibly lowering forest maturity in upper elevation settings, thereby improving ungulate forage and huckleberry habitat. [8]
This section assembles available data to model the late Pleistocene and Holocene climate sequence for the Mount Rainier region. Sources include plant macrofossil and pollen data and interpretive summaries from Mount Rainier, the Cascades and the surrounding region (especially Dunwiddie 1986, Heusser 1977; Whitlock 1992; and Sea and Whitlock 1995); and synchronous North Cascade and Mount Rainier glacial advance-retreat patterns (Crandell and Miller 1974; Porter 1976). Climatic implications of these data are summarized in Table 2.3. The curve is adapted from Heusser's (1977) inferences for the Pacific Slope of Washington as modified by pollen and geological data. The modified Heusser curve is a useful means of modeling climatic change because it displays greater variability and provides a better notion of climatic process than static time-block sequences (see Sea and Witlock 1995:378 for a good summary of the latter). Even though the curve simplifies a more complex natural reality by smoothing shorter-term oscillations, it can be refined as additional data become available and offers a more precise mechanism for comparing human system responses, if any.
The climate curve in Table 2.3 estimates long-term changes in summer temperature based on relative frequency of western hemlock (Tsuga heterophylla) to Douglas fir (Pseudotsuga menziesii) in 300 pollen spectra from the Hoh River Valley on the western Olympic Peninsula and Salmon Springs near Puyallup in the Puget Trough (Heusser 1977), as modified by macrofossil and glacial data noted above. Changes are expressed in degrees Celsius relative to present average July temperature at sea level (zero on the scale). The vertical guides are set arbitrarily at 1° C above and below the modern July mean. This zone approximates historically experienced (i.e., modern) climatic circumstances. Portions of the curve falling to the left (i.e., colder) of the -1° C line are classified by reference to their most commonly applied regional glacial or interglacial nomenclature. Sections falling within the ±1° guidelines are classed as modern interludes. The single significant part of the curve falling to the right of the +1° line (i.e., warmer) is the mid-Holocene xerothermic period labeled here the Hypsithermal Interval.
Table 2.3 Mount Rainier Region Late Pleistocene - Holocene Climatic Sequence
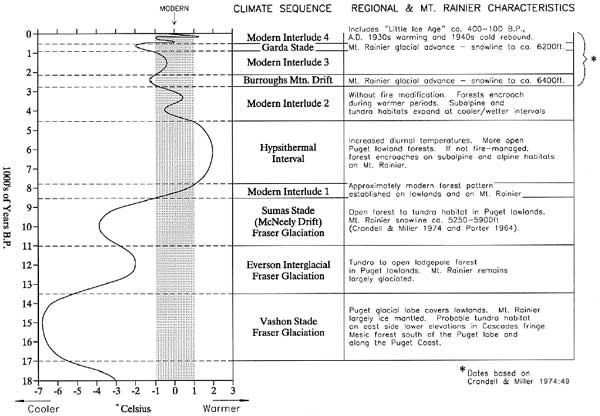
It now appears that Mount Rainier landscapes became sufficiently glacier-free, with adequate time for floral and faunal dispersal processes to have established economically useful plant and animal habitats by about 8,500 years ago. Prior to that time, glacial ice mass would have effectively precluded human use of Mount Rainier proper; although the Puget lowlands, eastern foothills and larger intermontane valleys could have sustained exploitable species as early as the Everson interglacial circa 13,500 years ago (the Pacific coast and southern lowlands earlier still). Assuming, for the moment, that humans did not intervene to alter natural successional processes, basic climate/environmental patterns on Mount Rainier and in the Pacific Northwest approximated the basic pattern shown in Table 2.3. The periods summarized below represent units of time during which the curve extended above or below 1° C of the modern July average. Please keep in mind that these periods are difficult to identify with precision and contain substantial climatic variability not represented in the curve's smoothed contours.
Late Fraser Glaciation: Vashon Stade; circa
17,000 to 13,500 B.P.
The Cordilleran ice sheet reached its maximum development in the Pacific Northwest between about 15,000 to 13,500 years ago (Whitlock 1992:9; Porter 1976:73; cf., Waitt and Thorson 1983). According to these sources, glacial ice covered the central and northern Puget Trough and the Straits of Juan de Fuca. East of the Cascades, Cordilleran ice lobes reached their most southerly extension. Alpine glaciers, however, appear to have been less extensive than earlier Fraser advances. Even so, Mount Rainier for all practical purposes was either ice shrouded or barren. Ice free areas on the mountain and in its immediate vicinity would have appeared essentially as rock or rock-grass islands impoverished of floral and faunal species in the midst of predominantly ice-mantled landscapes.
South of the glacial ice, vegetation took on a more mesic character compared with the preceding several thousand years. Probably responding to increased winter moisture that fed the Vashon stade, low elevation vegetation on the Washington coast, in the southern Puget Trough, and in lowland southwestern Washington shifted from predominantly parkland/tundra to parkland/open forest associations (Whitlock 1992:13-14). East of the Cascades and south of the glacial ice mass, periglacial steppe persisted to an elevation of about 1,500 m (4,900 ft).
Clearly, Mount Rainier, central and northern Puget Trough, Olympic Mountains, and North Cascades could not support human use during Vashon times. Even though plant and animal communities south of the glacial ice could potentially have supported human settlement, it is unlikely that anyone was there to avail themselves of them. At this time, most North American human populations remained locked in the ice-free periglacial Berginian refugium that extended from eastern Siberia to northwestern Alaska. It is unlikely that groups were able to move south of the continental ice in significant numbers (if at all) prior to its rapid wasting at the close of the Fraser/Wisconsin epoch (Burtchard 1987:209-213; Mathews 1982; Young 1982).
Terminal Fraser: Everson Interglacial &
Sumas Stade; circa 13,500 to 8,500 B.P.
Floral and faunal patterns changed dramatically with the wasting of cordilleran and continental ice at the end of the Fraser glacial epoch. Despite interruption by at least one major renewed advance about 11,000 to 9,500 years ago (the Sumas Stade, or McNeely drift on Mount Rainier), plant and animal communities dispersed into previously ice mantled areas, changing the character of the Cascades, Puget Trough and Olympic Peninsula dramatically. By virtue of its tolerance for nutrient poor, glacially scoured sediments, Pinus contorta (lodgepole) became the dominant tree species in central and northern Puget Trough early in the period (Whitlock 1992:15). Whitlock notes that through time "P. contorta was joined by Pica sitchensis [Sitka spruce], Pseudotsuga [Douglas fir], and Tsuga heterophylla [western hemlock] to form a more closed forest." Presumably these more nearly closed forest conditions appeared near the end of the period coincident with rapid wasting of the Sumas stade.
As Cascade alpine glaciers retreated, alpine tundra and patchy forest habitat established on lower mountains and intermontane valleys. Similar communities probably were established to a limited extent on Mount Rainier as well. During the McNeely drift, permanent snowline on Mount Rainier is estimated at about 5,900 ft with glacial tongues extending well into the major river valleys (Crandall and Miller 1974:43). At this elevation, many of the Park's larger mid-elevation ridge, cirque basin, and tarn parklands would have been under, or immediately adjacent to, perpetual snowpack. Vegetated habitat would have been limited primarily to low to mid-elevation slopes–probably in dispersed semi-isolated stands around the mountain base. While it is likely that some open forest/tundra species colonized the mountain during the Everson interglacial and persisted through the McNeely Drift (e.g., mountain goats, pika and marmots), their number was probably too low and exploitation cost too high to have been of serious economic interest to the few early Holocene human groups in the region. The probability of human forays into the immediate Mount Rainier area increases sharply, however, with rapid glacial retreat and establishment of more nearly modern plant and animal associations after about 9,000 to 8,500 years ago.
Modern Interlude 1; circa 8,500 to 7,800
B.P.
The demise of the Fraser glacial epoch, and ultimately the onset of the mid-Holocene hypsithermal interval probably was due largely to an increase in summer solar radiation brought about by more pronounced earth axial tilt in conjunction with a near sun orbit (perihelion) during the summer (Kutsback and Guetter 1986; Whitlock 1992:15-16). According to Sea and Whitlock (1995:379), "The direct effects of greater-than-present summer radiation in the Pacific Northwest included higher temperatures and decreased effective moisture. Increased summer radiation also resulted in a strengthening of the east Pacific subtropical high pressure system." In concert, these events induced dramatic environmental changes in the Pacific Northwest and over much of the world.
During this relatively brief interval, environmental conditions on Mount Rainier are likely to have approximated those of the present day–though perhaps with hotter summers. In essence, this is a formative period during which Mount Rainier took on most of its modern vegetative character; even though, geologically, the mountain was rather different than present. The mid-Holocene destruction and rebuilding of Mount Rainier's summit had yet to occur, and many of the newly ice-free landscapes are likely to have been highly unstable. Even though the permanent snowline retreated to approximately its present elevation, the effects of rapid glacial retreat should have lingered. It is likely that water-heavy ice masses stimulated frequent avalanches on upper elevation slopes and generated repeated high energy lahars in glacial river valleys. Newly exposed, poorly consolidated high gradient terrain should have been subject to repeated mass wasting rock avalanche movement.
These geological characteristics probably did not have a marked effect on forest succession above the lahar-prone river valleys and below glacial margins. It is likely that, for the first time, most of Mount Rainier's low to mid-elevation slopes took on a forest mantle roughly comparable to that of the present day, though probably with greater species variability and higher fraction of seral representatives. It is reasonable to expect basic faunal patterns described earlier in this chapter to have become established as suitable habitat became available.
The development of economically useful plant and animal associations and the presence of human populations sets the stage for initial use of Mount Rainier National Park. Even so, because of very low regional population density and probable availability of game in less logistically challenging places elsewhere, it is likely that use of the mountain was limited. Indeed, people may have done little more than observe the massive peak at a distance well beyond present Park boundaries.
Hypsithermal Interval; circa 7,800 to 4,500
B.P.
Because the northern hemisphere had entered into a period of rapid climatic change, the first Modern Interlude was essentially a transient state into a prolonged xerothermic period that dominated the region for over 3,000 years. Evidence for the existence of an early to mid-Holocene drought in the Pacific Northwest is compelling, though timing and local effects remain incompletely understood. Heusser (1977) notes its impact on the Puget Trough and coastal Olympic peninsula by charting an increased fraction of Douglas fir over western hemlock–a change that he believes reflects an increase in about 2° C in mean summer temperatures compared to the present climate. [9] Citing a number of sources, Whitlock (1992:17) reinforces the point, noting further that prairies and open forest conditions expanded throughout most of the Puget Trough. Fire frequency increased, in places reducing forest maturity in a manner that created a "mosaic of forest in various stages of succession." At Battle Ground Lake on the lower western flank of the southern Washington Cascades, Whitlock's pollen data suggest a 40-50% reduction in annual precipitation between 9,500 and 4,500 B.P. Similar to Heusser, she suggests that these data imply an annual temperature 1° to 3° C higher than present.
East of the Cascades, the forest/steppe margin moved higher and further north than present. The Columbia Plateau remained essentially treeless. Higher than present percentages of chenopodium and amaranthus at Carp Lake in southwest Washington indicate warm/dry rather than cold/dry condition were dominant during the period (Whitlock 1992:17; Barnowsky [Whitlock] 1985).
On Mount Rainier, Dunwiddie (1986) examined pollen and macrofossil data from three sites situated between 4,250 to 4,920 ft elevation on the mountain's southern flank atop the 6,000 year old Paradise Lahar. As expected for a very young landscape, Dunwiddie notes a high fraction of early seral species at the base of the profiles (especially noble fir [Abies procera], subalpine fir [A. Lasiocarpa] and lodgepole [Pinus contorta]). However, seral species continue to dominate all levels below Mt. St. Helens series Yn tephra (circa 4,000 to 3,250 B.P.), suggesting persistence of a warmer/drier than present climate before that date (Dunwiddie 1986:63). Assuming his interpretation is correct, these Mount Rainier data are approximately consistent with the general mid-Holocene xerothermic period as indicated by floral changes elsewhere in the Pacific Northwest. [10]
Glacial drift data from Mount Rainier also indicated a prolonged mid-Holocene period of alpine glacial withdrawal. No major glacial advance can be documented between the final retreat of the Fraser (McNeely Drift) circa 9,500 years ago and the relatively limited advance marked by the Burroughs Mountain Drift about 2,900 years ago (Crandell and Miller 1974). This does not mean that there were not lesser glacial advances and retreats during the period, but that such advances were small and traces obliterated by later, larger advances.
Geological events on Mount Rainier would also have affected environmental patterns temporarily, particularly to the mountain's southern and northeastern slopes. About 6,000 years ago, the Paradise and other lesser lahars reduced the summit and modified landforms on the south face of the mountain. Later, the 5,700 year old Osceola collapse and mudflow lowered the summit approximately 2,000 ft, moving a massive sediment load down the north and east faces of the mountain into the White River drainages and out into the Puget Trough. Undoubtedly, the physical character of the mountain was transformed dramatically. Even so, it is likely that impacts to vegetation patterns were largely temporary. The impact to the archaeological record notwithstanding, effect on land-use itself was probably neither substantial nor long lasting. The primary effect would most likely have been limited to short-term repression of forest cover serving to enhance somewhat the resource productivity and economic utility of affected landforms.
Clearly, regional effects of the Hypsithermal Interval are complex. Since human populations were now established in the region, Hypsithermal climatic changes hold implications for adjustments to regional settlement patterns and for use of Mount Rainier itself. For our purposes, the following general Hypsithermal patterns appear to apply to subregions in the vicinity of Mount Rainier:
East of the Cascades:
1) Increased aridity and degradation of ungulate forage in the Columbia Plateau.
2) Northward and upward shift in forest/steppe margins in the Okanagan and Cascade Mountains respectively.
West of the Cascades (Puget Trough):
1) Increased patchiness and open forest mosaics with increased fraction of mildly drought tolerant species such as oak (Quercus) and Douglas fir (Psuedotsuga).
2) Improved ungulate habitat due to opening of the forest canopy and concomitant expansion of grasses and brushy plant cover.
Southern Washington Cascades (including Mount Rainier):
1) On lower slopes and intermontane valleys, forest cover becomes patchier as drier conditions make forests more susceptible to fire–particularly in leeward settings; ungulate and collectable plant habitat improves.
2) Effect on upper to middle elevation slopes variable with exposure; species composition may have shifted to increased representation of drought tolerant species such as Psuedotsuga; in the absence of massive fire events, closed canopy forest should have dominated in wetter windward settings with increased patchiness in leeward settings and geologically disturbed landforms; effect on ungulate habitat unclear–here considered inconsequential.
3) Forest expansion on high elevation landscapes; forests invade and become more uniform in subalpine and lower alpine zones due to suppressed snowpack and lengthened growing season; upper elevation ungulate habitat is restricted; selective contexts for increased human induced burning to enhance habitat.
As the mid-Holocene drought deepened, it is likely that climatic events combined to reduce forage essential to support ungulate populations and their human predators east of the mountains, while simultaneously increasing them in the Puget Trough and lower elevation Cascade valleys and foothills. Accordingly, the Hypsithermal Interval may have witnessed a punctuated increase in use of landscapes and productive environmental zones in the vicinity of Mount Rainier. It is likely the use of the mountain itself increased during the period, perhaps involving fire modification of upper elevation landscapes.
Modern Interlude 2; circa 4,500 to 2,800
B.P.
Heusser's (1977) data suggest rapid amelioration of mid-Holocene warming starting about 5,000 years ago. By about 4,500 years ago, climatic conditions appear to have cooled to within 1° C of the present summer average (see Table 2.3). Given that climatic conditions were not perfectly stable or as regular as indicated on Table 2.3, the general return to cooler and moister circumstances should nonetheless have induced environmental regimes on Mount Rainier approximating those of the present day. Dunwiddie's (1986) data from Paradise lahar are consistent with a such a change. He suggests that cooler/moister conditions are indicated by increased fractions of mountain hemlock (T. mertensiana) and Alaska yellow cedar (Chamaecyparis nootkatensis) above Mt. St. Helens series Yn tephra. Because the mountain building Mount Rainier series C eruptions had not yet occurred, Rainier retained its lower, open to the east (but presumably now well vegetated) profile created by the Osceola collapse over a thousand years earlier.
Less xeric climatic conditions following the Hypsithermal Interval should have reversed regional environmental patterns. In the Puget Trough, forest cover returned more completely closed canopy. Forest maturity also increased on lower elevation Cascade valleys and slopes. Higher, forests retreated, reopening subalpine and alpine habitats on Mount Rainier. East of the mountains, forests again moved downslope, and Columbia Plateau rangeland improved.
It is plausible that loss of lowland ungulate habitat in the Puget Trough and lower western slope would have increased resource stress on growing regional populations now well established in the region. Such conditions would have established a selective context favoring intentional burning to maintain forests at a more productive, lower maturity state. On Mount Rainier, seasonal use should have focused on subalpine to alpine habitats and at the margins of naturally and culturally induced burns as discussed in the previous section.
Burroughs Mountain Glacial Advance; circa 2,800
to 2,100 B.P.
The Burroughs Mountain advance is the first stade of the Winthrop Creek glaciation on Mount Rainier (see Crandell and Miller 1974:44-50). It is indicated in the Park by a few remnant moraines, most of which have been crosscut by the later, larger Garda stade. At the peak of the Burroughs advance about 2,600 to 2,400 years ago, permanent snowpack is estimated to have shifted downslope to about 6,400 to 6,200 ft–400 to 600 ft lower than the mid-1960s snowline. In essence, permanent snowpack would have covered what is now the Park's alpine tundra as shown on Figures 2.10 through 2.13. Subalpine associations are likely to have shifted downslope to what are presently Mount Rainier's upper forested slopes.
The Burroughs glacial advance is roughly contemporaneous with the most recent mountain building episode on Mount Rainier (i.e., the Mount Rainier series C eruptions that rebuilt the summit to its present elevation. Series C events not only built the modern summit, but deposited sand to gravel sized tephra over much of the central and northeastern part of the Park (see Figure 2.3). It is possible that the combination of lowered snowline and heightened eruptive activity reduced the value of affected mid to upper elevation landscapes. More likely, however, increased snowpack, shortened growing season and periodic volcanic disruption simply pushed tundra and subalpine associations downslope, and perhaps extended them to the northeast. [11] It is unlikely that these events had marked effect on the productivity of economically useful plants and animals. Since major river valleys are not primary resource areas in any case, expansion of glaciers and glacial outwash events into them should not have significantly influenced human use.
Modern Interlude 3; circa 2,100 to 900
B.P.
Huesser's (1977) lowlands data and Crandell and Miller's (1974) glacial study indicate a circa 1,200 year interval between the two Winthrop Creek glacial advances. Huesser's climatic inferences suggest that temperatures rose overall during the interglacial period, but remained slightly cooler than present (see Table 2.3). During this interlude, regional floral and faunal patterns should once again have returned to near modern conditions as described for the second Modern Interlude above. On Paradise lahar, Dunwiddie's (1986) pollen profiles show an increase in western hemlock (T. heterophylla) about 2,000 years ago at Jay Bath, and Alaska yellow cedar (C. nootkatensis) at Reflection Pond about 1,500 years ago. Both suggest continuing cool and moist conditions, perhaps associated with natural succession processes begun with the amelioration of the Hypsithermal Interval.
Garda Stade Glacial Advance; circa 900 to 500
B.P.
The second major advance of the Winthrop Creek glaciation extended beyond and obliterated most of the moraines of the earlier Burroughs Mountain drift. The Garda stade on Mount Rainier reached its peak about 700 years ago, lowering permanent snowpack to about 6,200 ft–circa 800 ft lower than the mid-1960s (Crandell and Miller 1974:45). There is little floral data from Mount Rainier directly attributable to this period. Dunwiddie (1986:65-66) speculates that predominance mountain hemlock (T. mertensiana), normally represented at higher elevation, and loss of seral species at Jay Bath may be attributed to this glacial advance. 12]
In the absence of more refined floral data, we are limited to inferences based on combined effects of cooling and the lowered snowline. At 6,200 ft, permanent snowpack would cover some of the Park's larger expanses of upper elevation low gradient terrain (e.g., Sunrise Ridge and Spray Park). Even though tundra and subalpine associations would have shifted downslope, the substitution of sideslope habitat for the broader "pasture" of these rolling uplands, implies a loss of ungulate forage overall. However, other areas of flat to rolling terrain–Grand Park, Mist Park, Indian Henry's Hunting Ground–lie below the snowline as inferred and probably retained tundra, if not upper subalpine, qualities. For present purposes, I apply the same assumptions as those for the lesser Burroughs Mountain stade–that is, that net losses of upper elevation subalpine/tundra habitat probably were balanced by habitat increases at lower elevation. From a human standpoint, it is likely that use of the mountain continued, but use intensity may have decreased overall or shifted from higher landforms like Sunrise Ridge, upper Berkeley Park and Spray Park to lower elevation, low gradient landscapes such as Mist Park, Indian Henry's Hunting Ground, or presently forested lower elevation ground. It is important to note, however, that these assumptions are speculative in the absence of paleoecological and archaeological data from a wider selection of Park locations. Such work holds intriguing promise for improving our understanding of paleoclimate, habitat and human interactions and are highly encouraged.
Modern Interlude 4; circa 500 B.P. to
Present
The current interlude is marked by renewed climatic warming to within 1° C of the present July average at sea level. The period is associated with a variety of short-term, punctuated climate shifts world-wide (the apparent contrast with earlier periods simply reflecting more complete climatic data for the historic period). The maximum extension of glacial ice associated with the Garda stade retreated sharply at the outset, though renewed minor advances occurring between 500 and 100 years ago (i.e., the Little Ice Age) technically may be considered part of the general Garda event (Crandell and Miller 1974:49-50, also see footnote 12). For the most part, the period from A.D. 1450 to the latter half of the 1800s was cooler than present, but warmer than the Garda stade peak noted above. Twentieth century oscillations also include the sharp, but brief, xerothermic event of the 1930s and a cold rebound in the 1940s.
It is reasonable to assume that climatically sensitive environmental zonation patterns described earlier applied, in general terms, throughout this most recent interlude. Also we assume that resource opportunities described in that section were available to human populations during this time frame and during each of the earlier Holocene modern interludes. The manner and intensity with which they were exploited, however, varied not only with resource availability and abundance, but with population density and social variables affecting human groups in the broader region surrounding Mount Rainier. Consideration of broad-scale, long-term changes in regional land-use patterns and their implications for the archaeology of Mount Rainier National Park is the subject of the following final section of the chapter.
ENVIRONMENT, HUMAN USE AND MOUNT RAINIER'S ARCHAEOLOGICAL
RECORD
The intent of this chapter has been to draw attention not so much to Mount Rainier's environmental characteristics per se, but rather to emphasize their critical role in 1) conditioning the manner in which Park landscapes were used during the prehistoric past; and 2) transforming, obscuring, or obliterating the archaeological record of that use. The former issue largely concerns why, when, where and to a certain extent how the Park was used, and what effect such use can be expected to have had on the archeological record. The latter issue affects our ability to understand those land-use patterns through the physical remains left behind.
The second issue–post-depositional impacts to the archaeological record–is the more straightforward of the two. At Mount Rainier, the physical character of prehistoric remains have been altered or obscured most significantly through both destructive and depositional volcanic events and, perhaps, from major lahars. Lesser, more spatially localized disturbances result from such sediment disturbing events as tree fall, rodent burrowing, landslides and frost heaving. The primary concern here has been directed toward major impacts to the archaeological record resulting from Holocene volcanism–particularly Mount Rainier's summit collapse circa 4,700 years ago, and the summit rebuilding Mount Rainier series C eruptions of circa 2,400 years ago. The former event would have destroyed much of the early archeological record in the Park's northeast quadrant. The rebuilding eruptions served to hide what remained of the early to mid-Holocene record across much of the same quadrant as shown in Figure 2.3.
Important as physical alteration of the landscape may be to transforming the archaeological record, our primary interest here lies with the first of the two issues–the relationship between environmental patterns and prehistoric use of Mount Rainier. Accordingly, various sections of the chapter have sought to describe environmental parameters, and to draw attention to the causal relationship between geology, ecology and patterned human use of the mountain and its surrounding terrain. Below, issues developed in the text are summarized in terms of constraints Mount Rainier's environment posed to prehistoric human use.
Implicit to the discussion has been the notion that seasonal availability of economically important resources constitutes the primary reason that prehistoric humans used Mount Rainier. [13] I have argued that, because the most useful of these resources (e.g., goats, elk, deer, marmots, bears, game birds and huckleberries) are not evenly or randomly distributed, successful populations were those that fit their hunting and gathering strategies to the distribution patterns of the resources sought. In essence, this perspective views humans as intelligent predators focusing on a finite suite plant and animal species. If we hope to understand systematic human predation patterns then it is necessary for us to understand the behavior and patterned distribution of its most important prey. Once such links are established, corresponding patterns in the archaeological record can be predicted. Validity of the predictions become a test for the model and its economic premise.
Operating from these assumptions, and provided we can reasonably reconstruct environmental and resource history of a place, it is possible to derive basic expectations as to how people are likely to have organized their behavior in regard to that place. We have taken some time to establish the environmental resource history of Mount Rainier in order to build expectations that can be taken to the archaeological record for evaluation and rejection or refinement. Several basic sets of environmentally derived land-use and archaeological expectations close the chapter following the why, when, where and how metaphors introduced above.
Why Did Hunter-Gatherers Use Mount
Rainier?
Perhaps the most basic question regarding prehistoric use of Mount Rainier is why a seemingly remote, difficult-to-access place with severe winters would have been used by prehistoric people all. It was not long ago that high montane environments were assumed by many to be poorly suited to human use during prehistoric times; at least in other than the most cursory manner or as barriers to be crossed on the way to somewhere else. Ethnographic evidence to the contrary (Smith 1964), presumed unimportance of high elevation landscapes was implicit in the river valley emphasis of the Park's first formal archaeological survey (Daugherty 1963) and lingered into the mid-1970s as witnessed by statements alluding to the Park's limited potential for archaeological remains (Bohannon 1974, 1975).
To understand why mountains were important to prehistoric hunter-gatherers, we must recognize that such places were not particularly remote to early Holocene foragers whose mobility options were not restricted by ties to lowland villages. Most archaeologists now accept basic tenets of Binford's (1980) forager/collector model as modified and elaborated in the Pacific Northwest. [14] Working from that perspective, the annual round of early Holocene, small foraging groups would have been determined largely by the seasonal availability and productivity of locally obtainable subsistence and maintenance resources. Population density and competition for available food was low. The relatively few foraging groups in the region were free to move to new hunting and gathering places in a reasonably unencumbered fashion in accordance with changes in the availability of the most important resources. Storage needs were minimal and residential moves and could be made by the entire social unit.
For early Holocene foragers, subalpine and alpine habitats were used simply because they were among the best available places to acquire useful plants and animals during the summer and early autumn. Mountains remained useful so long as resource competition was low, and people were mobile enough to move into them during the productive summer season and move away to new locations when winter returned.
During the mid to late Holocene, the economic draw of montane habitats is less clear. Assuming increasing regional population density and elevated resource competition, unfettered use of upland habitats by small, autonomous groups would at some point have had to become unreliable. With or without environmental changes, a time had to come when there simply were too many people competing for too few resources in too little space. Such conditions provide economic stimulus for a shift to more intensive use of lowland resources–especially salmon; relying on mass harvest and storage (rather than group relocation) to bridge seasonal short-falls. Group sizes become larger and movement restricted by labor and storage requirements of the new economy. In essence, mobile foragers become semisedentary collectors with late summer harvest responsibilities focused on lowland rivers rather than subalpine meadows.
Schalk (1988) presents the clearest arguments for decreased mid to late Holocene use of upland environments in his study on the Evolution and Diversification of Native Land Use Systems on the Olympic Peninsula. As suggested above, he argues that salmon acquisition, processing and storage conflicts with summer use of the mountains, decreasing the resource value of the uplands and increasing logistical difficulty in their exploitation. Rather, it made better sense to allow elk and deer to fatten safely in subalpine summer habitats, to be hunted in the winter after they migrated back to lower elevation grazing areas.
The strength of these arguments not withstanding, presence of archaeological materials atop Mount Rainier-C tephra suggests that use of Mount Rainier's subalpine habitats continued into the late Holocene. [15] Three logical possibilities accommodate these data: 1) lowland-based communities continued to dispatch groups to the mountains to collect high-value commodities despite logistic difficulty and high labor costs relative to return; 2) the mid-Holocene shift to collector economies was not perfect, leaving marginal forager groups free to continue summer use of montane habitats despite reduced reliability; and/or 3) collector systems extended territorial boundaries into upland habitats and dispatched groups for limited collection tasks while guarding against summer poachers. In all cases, some use of the upland habitats continues, but in a manner consistent with increasingly complex logistic and political mechanisms required to fit elevated population density to a finite resource base.
In sum, there is ample resource-based reason to expect use of Mount Rainier to have begun early in the prehistoric period and to have continued, perhaps in a per capita reduced and redirected fashion, throughout the Holocene. The extant archaeological record amply demonstrates prehistoric use of Mount Rainier and the Cascades generally. If the arguments presented here have utility, we can expect continued archaeological research to demonstrate an extended temporal period for such use, and if the data are adequate, changing land-use strategies in the middle Holocene.
When Did Hunter-Gatherers Use the Mountain?
Temporal issues regrding use of Mount Rainier include seasonality, onset of earliest human activity and potential variation in use intensity through time. The first two of these issues are imbedded in the discussion above. Clearly, seasonal use of subalpine and higher landscapes on Mount Rainier is constrained primarily to summer and early autumn by severe winter weather and heavy snow cover. Generally, snowpack has melted and brows have greened sufficiently to support upward ungulate movement in June. The warmest, most predictable weather occurs in August and early September. Subalpine habitats can remain open into October. Accordingly, we would expect human use to have been biased toward these seasons during the prehistoric past much as it is in the present.
Human use of Mount Rainier and the Cascades probably began in the early Holocene coincident with retreat of Fraser glacial ice, dispersal of adequate exploitable plant and animal habitat, and presence of mobile human hunter-gatherers in the vicinity. Available paleoenvironmental data suggest that post-glacial open forest and tundra habitat was established in the Puget Lowlands and Columbia Plateau east of the Cascades as early as 13,500 years ago. With final wasting of the Sumas stade, similar habitat should have been established on Mount Rainier in sufficient area to support ungulate populations about 8,500 years ago. Human presence has also been established in the greater Pacific Northwest by this time. Accordingly, it is possible that earliest human use of the Park dates to approximately this period. The probability for significant human use of Mount Rainier rises sharply after circa 5,000 to 4,500 years ago as Hypsithermal climatic effects weakened and forest retreated to form the modern subalpine pattern.
Variation in human land-use intensity attributable to Holocene environmental causes is more difficult to predict. Events that can be expected to have affected exploitable plant and animal productivity include Holocene glacial episodes, volcanism, and forest encroachment during the Hypsithermal Interval. It is likely that the impact of both glacial and volcanic events was limited to short-term or localized habitat disruption with only marginal impact on human use. It is unlikely that either of these sources would have been sufficient to have caused a land-use hiatus of sufficient magnitude to be visible in the archaeological record. Rather, use patterns are expected to have shifted temporarily to remnant productive zones away from the source of disruption. Holocene glacial advances are expected to have lowered subalpine habitat several hundred feet in elevation. Given sufficient temporal and spatial control, the archaeological record may show a comparable downward shift during prolonged glacial advances at about 2,400 and again at about 600 years ago.
Where Did Montane Hunting and Gathering
Activities Take Place?
Much of the discussion in this chapter has been dedicated to developing ecological concepts that would allow us to predict which mountain landscapes, or more properly environmental zones, are expected to be associated with most frequent prehistoric human use. Building on arguments linking edible resource productivity to repressed forest maturity, I have argued that human use, and hence the archaeological record, should demonstrate a consistent bias toward upper forest, subalpine and alpine tundra environmental zones. These are the places in which the most critical floral and faunal species – especially mountain goats, elk, deer, marmots, bear, ptarmigan and huckleberries–tend to occur in greatest density. If these resources are the primary attractors to prehistoric human use of the mountain, then the archaeological record should display a corresponding distribution pattern. While fire and other sources of forest perturbation may open roughly comparable habitat at lower elevation, these areas tend to reforest quickly and, hence, are less likely to experience continued use. Accordingly, over the longterm, I expect most use to have concentrated at or near the more permanently immature subalpine to alpine habitats. Available archaeological site distribution data presented in Chapter 3 and visible in Figures 2.10 through 2.13 are consistent with these expectations.
The expected location of sites at the upper forest margin warrants special comment. Sites here could be explained either 1) by reference to retreating tree line during Holocene glacial episodes; and/or 2) because such locations minimized distance to subalpine hunting grounds in a relatively stormprotected setting. I suggest that forest margin sites may have served as base camps from which subalpine and alpine hunting and gathering forays were staged (cf., Baxter 1986). These upper midelevation residential camps may have been particularly desirable for mid to late Holocene collectors, logistically tethered to lowland villages; though their protected location may well have made them desirable places to early Holocene foragers as well. If these concepts have merit, we should find a higher fraction of larger, multi-functional residential site concentrations among the forest fringe sites when compared with those in the subalpine and alpine zones. At least three presently documented sites–FS 95-10 below Mt. Ararat, FS 90-01 on the southern flank of Sunrise Ridge, and FS 71-01 near Buck Lake–appear to meet these criteria.
Some economically important resources, of course, are found outside subalpine and alpine habitats. Lithic sources, for example, reflect exposure rather than habitat considerations. Local tool stone is important because it reduces the need to carry elaborate, and heavy, tool kits to the mountain. Several suitable exposures of micro and cryptocrystalline silicate stone (cherts and jaspers) are located on Mount Rainier. One, in a forested setting on Tum Tum mountain in the Park's southeast quadrant, is a documented lithic procurement site (FS 90-04). Another presently unverified outcrop is reported near Mt. Fremont (Dalle-Molle 1988). While not quarry sites per se, what appear to be locally procured flakes are documented at sites near Mother Mountain (FS 95-06) and Windy Gap (FS 90-03 and FS 95-07). Additional lithic sources undoubtedly exist on the mountain as well. If found in reasonable association with use areas or trails to such areas, we can expect a good chance of finding evidence of quarrying and primary lithic reduction activities at these places. The link to resource zones emphasized here cannot be discounted altogether, but is necessarily secondary to the location of the sources themselves.
It is likely that some prehistoric/early historic effort was given to collecting cedar bark in valleys and on Mount Rainier's wetter slopes. Fibrous cedar bark was used historically for making, among other things, expedient containers for huckleberry collection and transport (see Mack 1996). Western red cedar (Thuja plicata) is limited to low elevation wet forests, but Alaska yellow cedar (Chamaecyparis nootkatensis) grows higher on the nearer potential berry sources. A copse of peeled cedars has been reported in the Park adjacent to Shaw Creek (Carl Fabiani pers. com. 1995). Two additional peeled cedars have been reported on Mount Rainier's southern boundary (Liddle, pers. com. 1996). It is likely that more sites will be found as surveys continue. The location and age of culturally modified tree sites, of course will be limited by habitat (especially that of Alaska yellow cedar) and the survival age of the trees (several hundred years).
Pedestrian and equestrian trails provided ingress and egress from Mount Rainier's resource zones. Sections of a trail most plausibly linking Sunrise Ridge to the Yakama Nation east of the Park have been reported in the vicinity of site FS 90-01 (Jack Morrison and Carl Fabiani pers. com. 1995, and Roger Drake pers. com 1996). There is no doubt that a network of trails extended into and across Mount Rainier during the prehistoric past. Unfortunately, trails tend to be poorly represented as archaeological sites because, where not documented historically, they tend to be invisible to archaeological scrutiny. Even so, historical documentation and early maps (e.g., USGS 1915) may be consulted for additional insight into trail location in the more recent past.
Ceremonial sites may or may not be associated with subalpine and alpine habitats. Ceremonial use frequently is ascribed to stacked rock and talus pit features common to the Cascades (see Winthrop et al. 1995). Elsewhere, I have suggested that many, perhaps most, of these features functioned as hunting blinds whose locations were keyed to the spatial association of rocky scree slopes with broad visual exposures (Burtchard and Keeler 1991:86) and hence are explainable by reference to the general resource driven model employed here. The validity of these arguments notwithstanding, ritual use of Mount Rainier cannot be discounted altogether. As yet no stacked rock or other arguably ritual features have been identified in the Park. However, the probability that such sites will ultimately will be documented is high. At that time, renewed effort should be made to establish function and investigate spatial patterns.
How Did Regional Settlement Systems and Montane
Environments Affect Site Distribution Patterns?
Hunting and gathering patterns on Mount Rainier are constrained by the particular suite of resources available, their tendency to aggregate in upper elevation habitats, and their strictly seasonal availability. I have suggested that montane resources are integrated differently into the seasonal round of early Holocene foragers compared with mid to late Holocene collectors tethered to villages in major lowland river valleys. For foragers, we expect small, mixed age and sex residential groups to move as a unit to the near vicinity of the resources sought during the season in which they could be exploited most effectively. Foragers are expected to relocate when economic return degrades through local overexploitation or seasonal change. Collector based economies support higher population densities operating out of permanent to semi-permanent villages. Resource collection trips are made to and from the village with composition of the collecting group dictated by the character of the task.
In either case, once on Mount Rainier, ungulate and other faunal resources probably were acquired in a roughly similar manner. Because of exposed habitat, unpredictable weather and occasionally rugged terrain, upland game are most effectively hunted by small task-specific parties capable of rapid place to place movement with short-term stays at any given point. Since such characteristics are not common to mixed residential groups, it is possible that both foragers and collectors on Mount Rainier employed a two-part settlement pattern with hunting parties operating out of and returning to somewhat larger residential base camps–the latter perhaps situated at the forest fringe.
Whether collected and dried for return to lowland villages or foraged for immediate consumption, huckleberries were probably the single most important floral resource on Mount Rainier. Acquisition requirements for huckleberries are quite different than those for game animals. Traditionally, huckleberries are collected by predominantly women and children and returned to a central location for consumption and processing. For early Holocene foragers, the optimal strategy overall would be to locate a base camp in a setting intermediate to berrying and hunting grounds that allowed access to both. Assuming huckleberries were valuable enough to warrant mass harvest, transport and storage, mixed sex hunting and gathering groups probably continued to use Mount Rainier through the mid to late Holocene. A dual residential gathering and task-specific hunting strategy appears to be the most optimal means to access both resource classes simultaneously.
Despite very different ways in which montane resources were integrated into broader subsistence systems through time, the manner in which they were most effectively exploited on Mount Rainier may have resulted is similar archaeological signatures. That is, for both early Holocene foragers and later Holocene collectors, the best way to hunt and gather mountain resources was to establish moderate-term residential base camps at the forest/subalpine ecotone in support of taskspecific procurement locations in subalpine, alpine and perhaps low glacial settings. Although limited, present site distribution patterns on Mount Rainier are consistent with these expectations. As shown on the fold out maps and discussed in subsequent chapters, prehistoric sites are strongly oriented toward subalpine to alpine contexts. Most of these most sites appear to be low density, probably task-specific, lithic scatters. Larger, ostensibly more functionally diverse sites, are set at somewhat lower forest/subalpine ecotonal settings.
| <<< Previous | <<< Contents>>> | Next >>> |
archaeology/chap2.htm
Last Updated: 18-Oct-2004