|
FOSSIL BUTTE
The Geologic History of Fossil Butte National Monument and Fossil Basin NPS Occasional Paper No. 3 |

|
PALEONTOLOGY (continued)
Vertebrates
FISH. The single factor that has made Fossil Butte world famous is the occurrence of literally millions of beautifully preserved fossil fish. In one layer approximately 14 inches thick, the fish are nearly black and preserved in a nearly white shale. Most of the fish are perfectly preserved, retaining every detail of the skeleton and even undisturbed scales. Fish from Fossil Butte have been collected and sold since the 1870s and may be found on sales counters, in museums, and on living room walls throughout the world.
The fossil fish from the Green River Formation have a history of discovery stretching back to 1856. At that time Leidy (1856:256) described a fish given to him by a Dr. John Evans. Leidy called the fish Clupea humilis, a type of herring. Exactly where Dr. Evans obtained the specimen is not known. He passed through the area of the Green River Formation several times and could have collected it during any one of those trips (Knight 1955:12).
During the construction of the Union Pacific Railroad in the 1860s, workmen blasting a cut through Green River shales, about 2 miles west of Green River, Wyoming, came upon remains of well-preserved fish. Hayden (1871:742) first mentions this cut as "Petrified Fish Cut." Many of the fish from this cut were given to Cope who described them in Hayden's (1871) report. Insects, plants, and a bird feather were also obtained from "Petrified Fish Cut" at this time.
Sometime in the 1870s the first fish were obtained from Fossil Butte itself. Cope (1877, 1878) described fish from the Green River Formation which may be from Fossil Butte. His locality is "nearer the mainline of Wasatch Mountains" (Cope 1877:807). This may be Fossil Butte. His 1878 locality information is just as poor. He does mention a specimen of Priscacara peali given to him by A. C. Peale. Since it was Peale who first wrote about Fossil Butte, the specimen may have come from the butte.
Peale (1879) first mentioned the quarrying of fish at Fossil Butte, but he had no specific date as to the discovery of the butte or the beginning of quarrying of fish.
Cope (1884) published two large volumes about the Tertiary vertebrates of the West. Here he mentioned three fossil localities for his Green River Fish: "Petrified Fish Cut," "The Mouth of Labarge Creek," and "Twin Creek." Twin Creek is now known as the Fossil Butte site.
Since that time, the quarries at Fossil Butte have been extensively worked, mostly by commercial collectors.
The fish from Fossil Butte form an unusual array of genera (Table 1) unlike any now found living together. Included in the assemblage are forms usually found in marine waters and tropical, fresh-water fish (Schaeffer and Mangus 1965).
TABLE I. Simplified classification of fish from Fossil Basin
Elasmobranchii
Trygonidae
Xiphotrygon (sting ray)
Chondrostei
Polyodontidae
Polyodon (paddle fish)
Holostei
Lepisosteidae
Lepisosteus (gar)
Amiidae
Amia (bowfin)
Isospondyli
Osteoglossidae
Phareodus
Gonorhynchidae
Notogoneus (sand fish)
Clupeidae
Diplomystus (herrings, shad)
Knightia
Ostariophysi
Siluridae
Ameiurus (catfish)
Xenarchi
Aphredoderidae (pirate perches)
Erismatopterus
Asineopidae
Asineops
Acanthoptergii
Percidae
Mioplosus (perches)
Serronidae
Priscacara
More than 40 species of fish have been described from the Green River shales, but authorities do not agree on how many of these are valid. Because of this, only super specific groups will be considered in the following discussion.
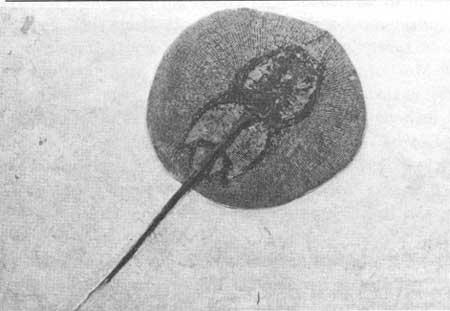
One of the strangest forms to come from Fossil Butte is that of Xiphotrygon (Fig. 14). This is not a true fish, but is a sting ray, related to sharks. Closely related forms still survive, with little change, in coastal waters in many parts of the world. Like sharks, sting rays have a skeleton composed of cartilage. Normally, cartilage is not preserved as a fossil as it disintegrates readily. The excellent skeletons of Xiphotrygon are good evidence of how well the shales of the Green River Formation preserve the fossils. Xiphotrygon is characterized by being shortened from back to front and flattened from top to bottom. It possesses a long, whip-like tail and bears enormous, flat pectoral (anterior) fins. The mouth is on the underside of the body indicating that Xiphotrygon, like its modern relatives, fed along the bottom of the lake. The strong flattened teeth form something like a pavement in the mouth, suggesting that it fed on clams and other hard-shelled invertebrates. Close relatives of Xiphotrygon live mostly in marine waters, but occasionally enter fresh water.
|
| Fig. 14. Xiphotrygon, a fossil ray. These forms are present but rather rare in the fish deposits of Fossil Butte. Length of original, 39 cm. Collection of University of Wyoming. |
The paddlefish, Polyodon, is rare but present at Fossil Butte. Similar forms still survive in China and the United States. The prominent features of Polyodon include a virtually scaleless body, along, depressed snout, and long gill rakers. The skeleton, unlike that of most modern fish, is composed of cartilage, hence skeletons of Polyodon are rare. The difference between the modern and Eocene paddlefish are so slight that the two probably filled similar or the same ecologic niches. This consists of feeding on plankton and other small organisms which can be obtained from the water. The long gill rakers would provide a large surface area on which food could be trapped as the water passed out through the gill slits. The large mouth would also provide a large surface area for catching and trapping food.
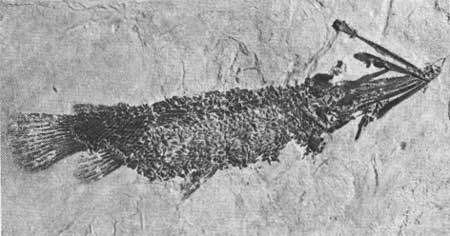
Quite common to streams and rivers of North America from the Cretaceous to the Recent is the garfish, Lepisosteus (Fig. 15). The most striking feature of the gar is its diamond-shaped scales, which are extremely hard and shiny. The body is long, essentially of the same depth throughout, and ending in a broad-based tail that is slightly asymmetrical. The mouth is armed with many small, sharp teeth for catching prey. One specimen over 5 ft long has been found in the Fossil Butte area.
|
| Fig. 15. The fossil gar Lepisosteus. Only a few gar have been found at Fossil Butte itself. Length of original, 70cm. Across the highway from the butte, one specimen over 5 ft long was found by Carl Ulrich. Collection of University of Wyoming. |
Fossils of the bowfin Amia (Fig. 16) are recorded from the Green River shales at Fossil Butte and in stream deposits from Cretaceous to Recent. This fish is still another "living fossil." Amia is a nocturnal predator. Presumably, the Eocene Amia had similar habits. The body in Amia has become elongate, as has the dorsal fin. The thickness of the scales has been reduced, but they are still rather heavy and cover the body. Specimens are rather large and deep-bodied. The tail fin is nearly symmetrical above and below. Amia is rare in the Fossil Butte fish quarries.

|
| Fig. 16. Amia, the bowfin. This too is rare but present at Fossil Butte. Length of original, 67 cm. Collection of American Museum of Natural History. |
By far the most abundant fish from Fossil Butte is the genus Knightia (Fig. 17, 18). This fish is a member of the Clupeidae, a family that includes modern herrings, shad, and sardines. The tail fin is small relative to the rest of the body. A row of modified scales extends on the back from the skull to the dorsal fin. Possibly they aided in streamlining the fish. Knightia apparently fed on the large amounts of plankton, especially algae, which lived in the waters of Fossil Lake. Fossils of Knightia are also known from Eocene lake sediments in South America.

|
| Fig. 17. The fossil herring, Knightia. This fish is by far the most abundant in the fish beds at Fossil Butte and at least in one layer was involved in a catastrophic mass mortality. Length of each fish, 13 cm. Collection of University of Wyoming. |

|
| Fig. 18. The herring, Knightia. |
Knightia appears to have been susceptible to mass mortalities. Some layers of shale at Fossil Butte contain literally hundreds of thousands of these fish that must have suffered catastrophic mass mortality. Possible reasons for the obvious sudden death of so many fish will be discussed in the section on paleoenvironment and taphonomy.
Another Eocene relative of the herring found at Fossil Butte is Diplomystus (Fig. 19, 20). Like Knightia, Diplomystus bears a row of modified scales on its back. Diplomystus is the second most abundant fish at Fossil Butte. The jaw in Diplomystus has a rather pronounced oblique angle to it. The deepest portion of the body is directly behind the gill region, with a continuous narrowing of the body toward the tail region. Diplomystus is known from Cretaceous rocks of Brazil and Syria and from Tertiary sediments in Brazil and West Africa (Schaeffer and Mangus 1965). Close relatives of Diplomystus now inhabit the waters off the coasts of Peru and eastern Australia.

|
| Fig. 19. Diplomystus, another type of herring, is the second most abundant fish at Fossil Butte. Length of original, 12cm. Collection of University of Wyoming. |

|
| Fig. 20. A large Diplomystus with four small bass, Priscacara. Length of Diplomystus, 45 cm. Collection of University of Wyoming. |
Phareodus (Fig. 21) belongs to a family of fish, the Osteoglossidae, which began as marine forms and has since become a fresh-water family. At present, they are restricted to tropical rivers and lakes of South America, Africa, and Australia. In the past they had a much greater distribution, as its occurrence at Fossil Butte indicates, including North America. It is a deep-bodied fish with a large head. The anal and dorsal fins are close to the tail fins. The many sharp teeth in the mouth attest to the carnivorous habits of Phareodus.

|
| Fig. 21. A large Phareodus, fairly common in the Fossil Butte fish beds. Length of original, 54 cm. Collection of University of Wyoming. |
The family Gonorhynchidae, the living sandfish of the Indo-Pacific area, is represented at Fossil Butte by Notogoneus (Fig. 22). Notogoneus is very long-bodied and quite slender. The body width does not vary much from head to tail, although narrowing does occur in the tail region. The tail fin is symmetrical, forming a good rudder. The anal, dorsal, pectoral, and pelvic fins are all small. The body is covered with small, elongate scales. Notogoneus was apparently a bottom feeder, living on small organisms picked off or out of the bottom of Fossil Lake.

|
| Fig. 22. Notogoneus, a long, slender bottom-feeder, occasionally found at Fossil Butte. Length of original, 56 cm. Collection of University of Wyoming. |
The Recent catfish, Ameiurus, has also been found at Fossil Butte in the Green River shales. The form of the Eocene catfish was very much like the Recent one. The habits of the Eocene catfish were probably like those of the modern Ameiurus, i.e., adaptations to a scavenger type existence.
Rather common in the sediments of Lake Gosiute, but not yet known from the Fossil Basin, is Erismatopterus, a member of the family Aphredederidae or pirate perches. This small, elongate fish has few distinguishing characters, the rounded front edge of the skull being most distinguishable.
Asineops is an Eocene relative of Erismatopterus and also one of the pirate perches. Asineops is, however, placed in a different family, the Asineopidae. Asineops, like Erismatopterus, is a rather plain-looking fish. The dorsal fin is long relative to the rest of the body. The body is slightly deeper behind the gills than elsewhere, giving Asineops a rather common appearance.
The Percidae, or perches, are represented at Fossil Butte by Mioplosus (Fig. 23). Mioplosus, as indicated by its well-developed teeth, was a carnivore that probably preyed on its piscine relatives in Fossil Lake. The large head blends in well with a strongly built, long body. The anal and dorsal fins are subequal in size and positioned opposite each other. The tail fin is large and fan-shaped. It is easily identified by the presence of two dorsal fins.

|
| Fig. 23. Mioplosus, a fairly common perch from Fossil Butte. Length of original, 30 cm. Collection of American Museum of Natural History. |
The one genus for which no close living relatives can be found is Priscacara (Fig. 24). The family Priscacaridae is thought to be related to the bass. The family is known only from the Eocene. The size of Priscacara is extremely variable. The body is deep and nearly round like that of the common sunfish, the head blending in well with the body and its contours. The most distinguishing feature of Priscacara is the series of strong, stout spines supporting the anal and dorsal fins. These spines may have acted as cut-waters for better swimming and/or they may have protected Priscacara from its more voracious relatives.

|
| Fig. 24. Priscarcara, a small bass that is common in the Fossil Butte fish beds. Length of original, 13 cm. Collection of University of Wyoming. |
AMPHIBIANS. To date, no amphibian fossils are known from Fossil Butte. They must have been present, as this class of animals is recorded from Devonian rocks more than 350 million years old and from rocks younger than those at Fossil Butte. Living amphibians are typified by frogs, toads, and salamanders. The lack of amphibian remains at Fossil Butte is more likely due to nondiscovery than nonexistence.
REPTILES. Reptile fossils are very abundant in the Green River and Wasatch formations. Also a jaw of the three-horned Triceratops is known from the Cretaceous part of the Evanston Formation.
Small lizards are normally represented only by jaw fragments and vertebrae. Their remains are most often found by a process of washing and screening of the sediments containing the fossils. Snakes are also represented in the Green River Formation. Remains of these animals are also often restricted to jaws and vertebrae. A complete skeleton of a fossil boa, Boavus idelmani, has been recovered from the Green River Formation and is the most complete fossil snake from North America (Schaeffer and Mangus 1965).
Turtles are among the most abundant reptile fossils. They are usually found as isolated shell plates, although many complete shells are known. The turtles Trionyx (Fig. 25), Emys, Baena, and Notomorpha are known from the Fossil Basin.

|
| Fig. 25. An aquatic turtle, Trionyx, from the shore-line deposits of Lake Gosiute. Several turtles has been found in the fish beds near Fossil Butte. |
The largest Eocene reptiles were the crocodiles (Fig. 26). The presence of these predaceous animals is further evidence for a prevailing warm and humid climate during the Eocene times since their tolerance ranges include only comparable conditions at the present time. A few incomplete specimens have been recovered from near the monument.

|
| Fig. 26. The skull of a large crocodile from hear the shoreline of Lake Gosiute. Several crocodile fragments have been found at Fossil Butte beds. Length of original, 75 cm. Collection of University of Wyoming. |
BIRDS. Birds, which first appeared in Jurassic time, are rare as fossils in rocks of any age because their skeletal remains are exceedingly fragile. Their fossilization requires special conditions because many of their bones are hollow as a weight-reducing adaptation for flight. These hollow bones are especially susceptible to breakage and poor preservation. Scattered remains of birds, mostly limb bones, are found in the Green River sediments. Feather imprints have also been recovered. A complete skeleton of Gallinuloides, a bird related to the South American fowl-like chachalaca, is one of the most significant finds from the Green River Formation. Recently, Brodkorb (1970) described the wings of puff birds similar to those now living in South America from near Fossil Butte.
In the adjacent Green River Basin to the east, paleontologists from the University of Wyoming have found nesting grounds of Eocene flamingos in slightly younger sediments of the Green River Formation. These "rookeries" have yielded many hundreds of fossil bones of this curious bird. Since these bird fossils were found proximal to Fossil Butte, they demonstrate the near subtropical nature of the environment in the Fossil Butte area in Green River times.
MAMMALS. The rocks of the Evanston, Wasatch, and Fowkes formations contain various fossilized remains of mammals which once inhabited the Fossil Basin.
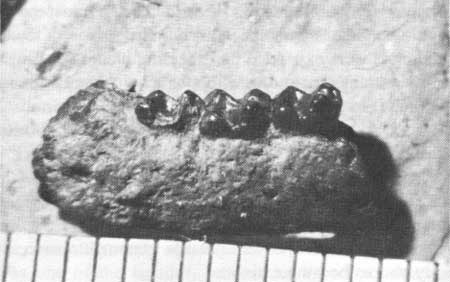
Mammal fossils are highly significant since they are used extensively for correlation and dating of the Tertiary rocks of the West. Fossils which are readily recognizable, limited in time, and widely distributed geographically are called "index fossils." These fossils are restricted to a particular time horizon in the rock. In this way, rocks of unknown age which are found to contain index fossils can be dated relatively and hence correlated with other fossil-bearing rocks. The small condylarth Haplomylus (Fig. 27) is a very early Eocene index fossil. Its presence in certain sediments in the Fossil Basin shows conclusively that these sediments were laid down in earliest Eocene times.
|
| Fig. 27. A jaw of the primitive mammal Haplomylus. Length of original, 1.3 cm. Collection of University of Wyoming. |
The mammal fossils found in Fossil Basin are also important in documenting the temporal changes in evolution and environment of the biotic community. The picture of Eocene Wyoming drawn through interpretation of these fossils is vastly different than that of the present day.
Mammals have a long evolutionary history that began over 200 million years ago in the Triassic period. At that time mammals had only just evolved from their reptilian ancestors. These earliest mammals were small, furtive creatures. However, with the extinction of the dinosaurs some 65 million years ago at the end of the Cretaceous, the mammals were able to diversify rapidly and fill the empty ecologic niches that the dinosaurs once occupied.
This filling of ecologic niches voided by the dinosaurs coupled with the spreading of mammals into hitherto unoccupied niches resulted in the development of a large variety of mammals. Some became extinct, others were the ancestors of modern mammals. Fossils of mammals found in sediments in the Fossil Basin are proof of the extent of this Late Cretaceous-early Tertiary mammal radiation.
Early mammals bore little resemblance to their later descendants. General trends in mammalian evolution were increase in size and the tendency to become more "modern"-looking through adaptation to changing Tertiary environments.
Most mammal fossils are extremely fragmentary. Teeth are most often preserved as they are the hardest part of the skeleton and therefore most resistant to wear and breakage. Thus, it often happens that the knowledge of a particular fossil mammal is derived entirely from its preserved dentition. This has obvious limitations for the completeness of our understanding about mammals in question.
The Paleocene portion of the Evanston Formation has yielded mammals of Torrejonian and Tiffanian age (Fig. 6).
The Torrejonian assemblage (middle Paleocene) (Gazin 1969) is small and poorly preserved. Its main significance is that the fauna records a definite time interval for the Evanston Formation. Gazin's work has demonstrated this fauna lobe intermediate in composition between similarly aged faunas recovered from sites to the north and south.
The major faunal elements are insectivores, primates, condylarths (primitive ungulates), and multituberculates (a type of extinct rodent-like mammal, with no known descendants).
The poorly preserved condition of these fossils makes discussion impossible except to note that they extend the known time range of mammalian habitation in the Fossil Basin.
The younger Tiffanian fauna (late Paleocene) (Gazin 1956) is also known from largely fragmentary remains. The multituberculate mammal Ptilodus, also present in Torrejonian deposits, is the most "primitive" mammal in the fauna. The multituberculates first appeared in the Jurassic (Fig. 4), and are therefore the oldest lineage of mammals that survived into Tertiary times. The molar teeth of these animals are characterized by multiple cusps arranged in parallel rows. Ptilodus, like most other multituberculates, was specialized in that its fourth lower premolars were expanded into shearing blades, the function of which is not known. Ptilodus was in many respects similar to rodents in the development of procumbant incisors and in general build and appearance. Multituberculates are, however, not related to rodents except that both are mammals. The differentiation of rodents in the late Paleocene and their diversification during the Eocene probably presented strong competition for the multituberculates who were unable to compete success fully and so became extinct.
The primates were represented by Plesiadapsis. This animal was about the size of a squirrel and had chisel-like incisors. Flesh reconstructions of Plesiadapsis, based on skeletal remains, show this primate to look much like a rodent with a long-snouted skull, clawed feet, long body, and tail. There is conceivable relationship between the rodents and Plesiadapsis-like primates.
The dominant Tiffanian herbivorous mammals were members of the Order Condylarthra. These primitive ungulates were diverse in both size and appearance. It is probable that later modern ungulates (Artiodactyla, Perissodactyla) evolved from the Condylarthra; however, there are no surviving members of this group. The Paleocene condylarths were mostly small animals and include such forms as Haplaletes, Litomylus, and Gidleyina. These mammals had somewhat insectivore-like teeth, and may have bridged the gap between insectivores and archaic hoofed mammals. One of the larger Tiffanian condylarths was Phenacodus. This mammal had along, massive skull and a long, probably flexible body. The limbs were stout and short. The body ended in a long tail. Small hooves were present on all digits, which numbered five per foot. The length of the largest Phenacodus was about 6 ft.
Carnivorous and omnivorous condylarthra were also common. Thryplacodon and Claenodon are two types known from the Evanston Formation in the Fossil Basin.
The true carnivores (Order Carnivora) were also present. The Paleocene carnivores were small and possibly arboreal in habit. The stem carnivore stock was represented by the family Miacidae. In the Fossil Basin the miacid Didymictis was common. In Didymictis, the mouth was armed with small, sharp teeth. Most important, the typical carnivoran carnassial (shearing) teeth had developed. Among carnivores, from Paleocene to Recent, the shearing blades developed on the last upper premolar and the first lower molars. Various kinds of shearing teeth have evolved in other mammal lineages.
Mammals from the Wasatch Formation in Fossil Basin represent the Greybullian and Lysitean provincial ages (see Fig. 6). These fossils are of earliest Eocene and mid to early Eocene age. Fossil localities in the Wasatch Formation of southwestern Wyoming are Knight Station (partly Lysitean), at Elk Mountain (Greybullian), and at Fossil Butte itself. The age of the Wasatch Formation at Fossil Butte is Lysitean.
The insectivores are represented by Diacodon, a small mammal known only from fragmentary material.
Primates also continued to expand and diversify in Wasatchian time. Microsyops was a very unusual type that belongs to the extinct family Microsyopidae. The teeth were sharp and adapted for eating fruit and/or insects. Microsyops was about the size of a rat or slightly larger. The microsyopid primates were limited to the Paleocene and Eocene and left no later known descendants.
Another group of primates is represented by Pelycodus. This was a small lemur-like animal that may possibly have been an ancestor of higher primates (monkeys, apes, and men). Pelycodus, like Microsyops, was an arboreal (tree-dwelling) animal which inhabited the forests of the Fossil Basin during the early Eocene.
The Order Taeniodonta is a strange and little known group of mammals. In the sediments at Fossil Butte the taeniodonts are represented by Ectoganus. This was a moderately large animal. The skeletal adaptations were similar to those of moles, but there was no actual relationship between the two groups. The front feet and legs were robust, and bore large claws. Ectoganus probably used these structures to grub for food, possibly roots. The incisor teeth were rootless and persistently growing. The tooth enamel was restricted to two bands on either side of the teeth. The single pair of upper and lower incisors was greatly enlarged. The taeniodonts became extinct in the Eocene and left no descendants.
Equally as strange a group of animals as the taeniodonts are mammals in the Order Tillodonta. These were herbivorous animals, some of which became quite large. They have many rodent-like characters, but were not related to rodents. In many respects, the morphology of rodents seems to have lent itself to convergence. Esthonyx was the most common tillodont from the Fossil Basin. Prominent, rootless incisors were a characteristic of the later members of the group. These incisors were chisel-like, as in rodents. The molars were unusual and bore certain resemblances to primitive carnivoran and insectivoran teeth. The tillodonts, however, bore uncertain relationships to other mammals and left no descendants beyond the middle Eocene.
In comparison with recent faunas, Paleocene rodents were rare faunal elements. Two early rodents were found in the Fossil Basin Wasatch Formation:
Paramys was relatively unspecialized but was diverse in the number of species. One species of Paramys was about 2 ft long and possessed a long tail. The body was long and slender. The skull was small and rather squirrel-like. The cheek teeth were quadrate and had low, blunt cusps. The typical, single pair of chisel incisors was present both on lower and upper jaws. Paramys was probably an arboreal form that did not look too unlike a modern squirrel and may have had a similar mode of life. Another rodent, Reithroparamys, was somewhat similar to Paramys in size and appearance. The differences between the two animals were mainly in the teeth and hind limbs. The latter showed certain very minor structural modifications that suggested a saltatorial (jumping) mode of locomotion for Reithroparamys.
Bats (Order Chiroptera) are extremely rare as fossils because their volant and cave-dwelling habitat as well as a fragile skeleton did not lend their remains to preservation. From the Green River Shales, from near Fossil Butte, a complete articulated skeleton of a bat has been found (Jepsen 1966). This animal, named Caronycteris index, was small and generalized in form. Superficially, it looked much like a typical brown bat. This fossil is important because it demonstrates that the bats had already become good fliers by early Eocene time.
The condylarths were prominent in the early Eocene but diminished in importance following this time. Phenacodus was still extant; however, it was replaced in importance by the smaller Hyopsodus.
Hyopsodus is perhaps the most commonly recovered Eocene fossil mammal. This animal was small and long-bodied and retained a more-or-less generalized structural pattern. In some respects Hyopsodus was similar to both insectivores and primates and was at one time or another regarded as belonging to either of these two orders.
Some of the later condylarths paralleled the more advanced ungulates. Indeed, the Perissodactyla (horses, tapirs, rhinos) and Artiodactyla (bovids, deer, pigs, sheep, etc.) were derived from early Tertiary condylarths. Meniscotherium was one such advanced condylarth. The cusps on its molars, instead of being blunt points, had developed into crescentic patterns, somewhat like those in deer or camels. The relationship of Meniscotherium to modern ungulates is only one of parallel dental development. Meniscotherium was a medium-sized animal, but about the same build and size as that of a cocker spaniel. Meniscotherium, however, had hoofs on its toes and was a forest-living browser.
The condylarth Haplomylus is found at the Elk Mountain Wasatchian locality. This small animal is typical of the Greybullian level of faunal organization (earliest Eocene). A supposed carnivorous condylarth was Pachyaena. Pachyaena was a fairly large animal for its time. The strong, robust jaws and teeth and heavy build suggest that Pachyaena was a predatory animal suited for preying on larger animals such as the amphibious pantodont Coryphodon. The affinities of Pachyaena to the condylarths are uncertain and are based primarily on foot structure.
The miacid carnivores were abundant and varied. Didymictis persisted and both Vassacyon and Vulpavus continued the arboreal, forest-dwelling habits of their Paleocene precursors.
The main carnivorous animals of the Eocene, however, were the creodonts (Order Creodonta) which were essentially early experimenters in carnivorous habits. As such they were diversely specialized. Proviverra (=Sinopa) was one of the smaller, predaceous creodonts that inhabited Fossil Basin during Eocene times.
One of the most characteristic fossil mammals from the early Eocene of Fossil Basin was Coryphodon. A specimen of Coryphodon was also the first mammal fossil lobe found in the Fossil Basin. The order of mammals to which Coryphodon belongs, the Pantodonta, was a group of large and heavily built, herbivorous mammals. Coryphodon was about the size of a small rhinoceros. The skull was large and heavily built. The brain, however, was small. The canine teeth were somewhat enlarged and strong. Stout limbs supported the bulky body. In habits, Coryphodon was possibly semi-aquatic.
The modern ungulates, Order Perissodactyla (odd-toed ungulates) and Order Artiodactyla (even-toed ungulates), first appeared in the early Eocene. The earliest members of both orders were small and relatively unspecialized.
The earliest perissodactyls were the horses and tapirs. The earliest horses were represented by Hyracotherium. This small, slender, and lightly built horse was about the size of a fox terrier. The front feet had four toes, and the hind feet three toes. The teeth suggest that this animal was probably a browser, living in the forests. In the Fossil Basin, tapirs were known from deposits of Lysitean (mid to early Eocene) age of which Heptodon was representative. The tapirs now live in the tropics of Malaysia and Latin America. Their present habitat suggests a similar environment may have been favored by the Eocene tapirs of the Fossil Basin.
The first Artiodactyls were pig-like forms, although they are only distantly related to true suids. Protodichobune was small and may have looked much like some of the early Perissodactyls. There are, however, a complex of structural features which serve to distinguish these mammalian orders from each other.
The mammal fauna from the Fowkes Formation places a late Bridgerian age on the Sillem and Bulldog Hollow members (Nelson 1973). Small mammals were most abundant in the Fowkes, especially primates and rodents. The marsupials were characterized by Peratherium which was a small, probably arboreal, opposum-like animal. Peratherium was but one stage in a slow and relatively conservative evolution of opposums that began in the Cretaceous.
The condylarths decreased in importance during Fowkes time and were gradually replaced by more advanced ungulates, the artodactyals and periossodactyls. Only the small Hyopsodus survived into the Bridgerian of the Fossil Basin. The horses continued to differentiate as evidenced by the appearance of Orohippus, an animal similar to Hyracotherium, but with minor dental differences. The primitive tapirs were represented by Hyrachyus, a medium-sized herbivore which was nearly as closely related to the rhinoceroses as it was to the tapirs.
The primates were also diverse in Fowkes time; however, this group began to decline in importance following Bridgerian times. As a group, the primates were restricted mainly to warm and forested environments. The lemur-like Notharctus and the tarsier-like Omomys and Hemiacodon were common in the Fowkes. Hemiacodon was typical of the late Bridgerian (an index fossil).
The true carnivores, represented by Miacis, remained small in size, perhaps to avoid competition with the larger surviving creodonts.
Insectivores from the Fowkes Formation are abundant and varied. Unfortunately, they are known almost exclusively from teeth. Fowkes insectivores include the hedge-hog-like Nyctitherium, Talpavus, and Scenopagus. Apatemys is an insectivore of uncertain affinities. The feeding habits of these early insectivores were probably similar to those of contemporary insectivore species (moles and shrews); however, there is some evidence to suggest many of these small creatures may have been semi-arboreal.
One of the most unusual of middle Eocene creatures is the giant Uintatherium. This was a large browsing animal, about the size of a rhinoceros. What was most unusual about Uintatherium was its skull which was large and strongly built. The large upper canines were apparently formidable defensive weapons. The skulls of the males bore six prominent bony protruberances which grew from the frontal region of the skull. The function of these structures is unknown. They may have been of use for defense.
During Bridgerian times, the rodents underwent an explosive adaptive radiation which ultimately may have led to the near extinction of the less well-adapted multituberculates. The diversity of Bridgerian rodents from Fossil Basin is impressive. Species of Leptotomus, Paramys, Thisbemys, Microparamys, Sciuravus, Mysops, and Pauromys demonstrate that a wide range of adaptations of environments was exploited by the rodents. This diversity in life habits has been a mark of rodent evolution from the middle Eocene to the present day.
The above descriptions of the known mammal fossils from the Fossil Butte area are intended to demonstrate the varied life forms that once existed in southwestern Wyoming. Further collecting will almost certainly expand the number of fossil mammal species from Fossil Basin and increase our knowledge of the succession of ancient environments which prevailed at different times in the Fossin Basin area.
TABLE 2. Mammals known from Fossil Basin
|
Faunal lists—
|
|
|
| <<< Previous | <<< Contents >>> | Next >>> |
sec4b.htm
Last Updated: 01-Mar-2005