|
YELLOWSTONE
The Bison of Yellowstone National Park NPS Scientific Monograph No. 1 |

|
CHAPTER 5:
POPULATION CHARACTERISTICS
THIS CHAPTER DISCUSSES THE REPRODUCTION and population structure and dynamics of Yellowstone bison during the period 1964-68. A series of bison reductions from 1961 through 1965 consistently removed a large number of breeding-age females. Records indicate that the reductions by field shooting of the mid-1950s also did this, as did some of the earlier removals at the Buffalo Ranch in Lamar (Table 9). Some population characteristics may change after a period without reductions. Data for all categories were not obtainable each year; hence, selected figures are used in some instances to suggest population conditions.
TABLE 9. Number of cows and calves removed during reductions.
| Year | Location | Cows | Calves | bTotal no. classified |
Percent of reduction | ||
| No. | Percent of total classified |
No. | Percent of total classified | ||||
| 1931-32 | Lamar | 184 | 90 | 1 | 205 | 92 | |
| 1932-33 | Lamar | 90 | 45 | 0 | 199 | 96 | |
| 1933-34 | Lamar | 59 | 85 | 0 | 69 | 39 | |
| 1938-39 | Lamar | 9 | 13 | 0 | 67 | 100 | |
| 1940-41 | Lamar | 110 | 54 | 0 | 205 | 100 | |
| 1941-42 | Lamar | 78 | 39 | 41 | 21 | 200 | 100 |
| 1943-44 | Lamar | 219 | 55 | 90 | 22 | 400 | 99 |
| 1945-46 | Lamar | 94 | 47 | 20 | 10 | 200 | 100 |
| 1949-50 | Lamar | 129 | 57 | 39 | 17 | 228 | 100 |
| 1953-54 | Lamar | 34 | 45 | 13 | 17 | 75 | 100 |
| 1955-56 | Lamar aMary Mountain bPelican |
15 123 53 | 62 53 45 |
2 59 32 | 8 26 27 | 24 231 118 |
100 100 100 |
| 1956-57 | Lamar aMary Mountain | 29 93 |
49 44 | 13 43 |
22 20 | 59 212 | 100 100 |
| 1961-62 | Lamar | 74 | 52 | 33 | 23 | 143 | 100 |
| 1962-63 | Mary Mountain | 187 | 52 | 101 | 28 | 357 | 100 |
| 1964-65 | Lamar Mary Mountain aPelican |
42 119 14 | 51 50 41 |
6 39 8 | 7 12 24 |
82 238 34 | 100 100 100 |
| 1965-66 | Mary Mountain | 18 | 36 | 9 | 18 | 50 | 100 |
bTotal classified (bulls, steers, calves) not necessarily total reduction number.
Sexual Maturity
Incidence of pregnancy by age class for two Yellowstone studies 25 years apart are compared in Table 10 as an indication of sexual maturity in the female population. The 1940-41 study sampled the semiranched population of Lamar; the 1964-66 study sampled the entire free-ranging population. The rates of pregnancy in the younger age classes are suggestive only, due to limited sampling. The 1940-41 study of the semidomestic herd indicated that an occasional female bred as a yearling. None (of 15 sampled) was found to be pregnant in the later study of wild bison. The 1964-66 study indicated that a few more bred as 2-year-olds and that approximately a fourth of the 3-year-olds were breeding. Both studies showed that the majority of females attained sexual maturity when 4 years old.
TABLE 10. Comparison of female pregnancy rates, by age classes, 1940-41 and 1954-66.
| Age class | No. sampled | Percent pregnant | ||
| 1940-41a | 1964-66 | 1940-41a | 1964-66 | |
| 2.5 | 2 | 7 | 0 | 14 |
| 3.5 | 4 | 11 | 50 | 27 |
| 4.5 | 13 | 7 | 92 | 71 |
| Young adult | 10 | 6 | 100 | 50 |
| Adult | 49 | 23 | 86 | 57 |
| Aged | 8 | 17 | 100 | 75 |
| Entire sample | 86 | 71 | 86 | 52 |
aFrom Skinner, Curtis K. 1941. Special report on Yellowstone National Park bison.
Yell. Natl. Park Bio. Files, 715-03. Buffalo (General). Typed.
| ||||
Sexual maturity for the female population as a whole may be occurring somewhat later now. The 1964-66 study of wild bison showed fewer females pregnant in the 3.5 and 4.5 age classes (27 and 71%, respectively) compared with 1940-41 study of the semidomesticated bison (50 and 929, respectively).
Sexual maturation also may be later in the Yellowstone population of this study compared with herds elsewhere. At Wood Buffalo National Park, 36% (160 sampled) of the Hay Camp and 59% (17 sampled) of the Lake Claire 2-year-olds were pregnant. Rates were 52% at both locations (92 and 31 sampled, respectively) for 3-year-olds (Fuller 1962). At Wichita, 74% (35 sampled) of the 3-year-old cows had calves (Halloran 1968).
Sexual maturity of males was not studied in Yellowstone. However, Fuller (1962) found that examination for spermatogenesis showed the age for males to be comparable to that for females: a few mature as yearlings, a third as 2-year-olds, and probably all others by 3 years of age. Halloran (1968) found that two experimental bulls were effective sires as 2-year-olds. According to observations made by McHugh (1958) in Hayden Valley in Yellowstone, males 8 years and older were the most active sexually, as indicated by "tending bonds"—the term he used for the relationship between a bull and a particular cow during the rut. Fuller (1960) also observed that only fully mature males tended cows. Apparently, sexual maturity in males occurs well before they are a part of the active breeding population.
Reproduction Rate
The present wild population showed a lower pregnancy rate in all age classes over 2.5 years compared with the semidomesticated herd of 1940-41 (Table 10). Females 3.5 years of age and older averaged 56 and 90%, respectively. Females 4.5 years and older averaged 62 and 90%, respectively.
Table 11 compares reproductive data recorded at Lamar by Rush (1932a), several later unpublished compilations also made at Lamar, and this study of the entire population. The pregnancy rate of 52% for females 2.5 years of age and older determined in this study was the lowest recorded in Yellowstone. The reason for this was not clear, because the age classes represented in all previous compilations (except 1940-41) were not recorded. The rate of 65% in 1931-32, which was lower than others prior to this study, may reflect a high percentage of aged females in that sample, as few cows were removed from the semidomestic herd before 1930. The influence of ranching practices such as winter feeding and culling of aged animals for many years may account for subsequent high pregnancy rates.
TABLE 11. Summary of reproductive data and fetal sex rations, 1931-32 through 1964-66.
| Year | Source | Number sampled | Total pregnant percent |
Pregnant and lactating percent | Pregnant only percent |
Lactating only percent | Number sampled |
Fetal sex ratio male/female | Percent unclassified |
| 1931-32 | Rush, 1932a | 184 | 65 | 52 | 12 | 24 | 54 | 108/100 | - |
| 1940-41 | Skinner, 1941 | 86 | 86 | - | - | 5 | 74 | 163/100 | 1 |
| 1943-44 | Yell. Natl. Park files | 219 | 75 | - | - | - | - | - | - |
| 1945-46 | Yell. Natl. Park files | 82a | 79 | - | - | - | 62 | 112/100 | 5 |
| 1949-50 | Yell. Natl. Park files | 115 | 94 | - | - | - | 104 | 138/100 | 1 |
| 1964-66 | Present Studyb | 71 | 52 | 3 | 50 | 1 | - | - | - |
aSample included only females 4 years of age and older. Other samples
included some females in the younger age classes.
bThis study sampled the entire population. Previous studies sampled only Lamar. | |||||||||
Yellowstone's present wild bison population showed a reproductive rate equal to the 52% reported by Halloran (1968), and lower than the rate among all populations compared by Fuller (1962). Halloran attributed the low rate at Wichita Mountains Wildlife Refuge to high calf survival. Calves were known to nurse into their second year of life, thus prolonging the physical drain on the cows (and perhaps influencing the possibility of conception). Fuller considered brucellosis infection and severe climate as possible adverse influences on reproductive rates in Wood Buffalo National Park.
Calf survival apparently was not an important influence on the reproductive rate determined by the present study. Although survival appeared high (see Calf Mortality), no yearlings were observed nursing during the study. Only 4% of the cows (Table 11) examined during the mid-winter reductions were still lactating.
Brucellosis in Yellowstone animals did not appear to be a factor in the low incidence of pregnancy. Rate of infection according to tests from 1964-66 averaged about 54% for Lamar, 42% for Pelican, and 26% at Nez Perce Creek in the Firehole area (Barmore 1968). These rates were all lower than the 62% at Lamar in 1941 when the pregnancy rate, at 86%, was much higher than now. Although Rush (1932a) mentions that a number of abortions were known to have occurred, abortions were apparently rare except in the crowded corral at the Buffalo Ranch (Dave Pierson 1968 pers. comm.). Quortrup (1945) stated that brucellosis had little or no effect on the herds, as did Tunnicliff and Marsh (1935) and McKenney (Skinner 1941).
Differences in population age structure, as reflected in the samples, partly explain differences in reproductive rates between the wild bison populations of Wood Buffalo and Yellowstone National parks. Subadults and young adults formed a much larger part of the Wood Buffalo National Park samples (Fuller 1962); aged animals formed more of the Yellowstone sample. However, all age classes (except the aged) in Yellowstone showed a lower reproductive rate.
The low reproductive rate in Yellowstone may result from a complex of environmental factors, in part related to severity of the winters. Both Yellowstone and Wood Buffalo National parks have severe winter conditions, but the details differ (Table 12). Fuller (1962) believed extreme cold, per se, had little effect on bison, but was important when there was wind. Wood Buffalo National Park, compared with Yellowstone, is colder, but has less snow and fewer periods of crusting conditions during the winter. Trees provide protection from the wind. Differences in amount of wind were not as apparent. Yellowstone apparently has a more severe winter environment, for bison, than does Wood Buffalo National Park.
TABLE 12. Comparison of winter climatic factors in Wood Buffalo and Yellowstone National Parks.
| Cold | Snowfall | Crusting | Wind | Wind protection | |
| Wood Buffalo National Parka | January mean daily maximum -7.8°F | 53" | apparently rare | rarely over 15 mph in extreme cold | woods everywhere interspersed with grazing areas |
| Yellowstone National Parkb | January mean daily maximum approx. 22°F | approx 150" | most winters, one or more periods | a winter average of more than 8.5 mph | no interspersed woods (except the Firehole), winter vallyes large and open |
aInformation from Fuller, W. A., 1962. The biology and management
of the Bison of Wood Buffalo National Park. Can. Wildl. Serv. Wildl.
Manage. Bull. Ser. 1, No. 16, 52 p.
bInformation from U.S. Dep. Commer. Climatological Summary for Yellowstone National Park (1930-59). Temperatures and snowfall are for the park interior. | |||||
The effects of the Yellowstone winters on subadults is suggested by the mortality among them, as discussed later. Winter conditions, perhaps through nutritional influences, may delay sexual maturity among subadults. Houston (1968) suggested that animal nutrition during winter periods might control yearling moose (Alces alces) ovulation rates. Robinette et al. (1955) attributed reproductive rates among yearling mule deer (Odocoileus hemionus) to the type of winter preceding that during which the pregnancy rates were determined. Possible variation in effects between mild and severe Yellowstone winters was not obtained during this study.
The influence of Yellowstone winters on reproductive rates among adult bison was unknown. Pregnancy rates among adult moose in Jackson Hole, south of Yellowstone, approximated those from other ranges (Houston 1968). Other environmental factors which were not apparent from this comparison of Yellowstone and Wood Buffalo National parks may be important in affecting adult reproductive rates.
Fetal Sex Ratio
The fetal sex ratio was not estimated during this study. However, records kept by Rush (1932a) and during four later reductions in Lamar (Table 11) showed more males among fetuses, ranging from 108 to 163 males/100 females and averaging 127 males/100 females (294 fetuses). This was slightly higher than the 112 males/100 females reported by Fuller (1962).
Number of Young at Birth
No twin fetuses were noted in any of the above records. One instance of twins in the Lamar herd was reported (McHugh 958). Other workers (Fuller 1962; Halloran 1968) and historical accounts agree that twins are rare.
Population Structure
Sex and age structure was suggested mainly by trap records made during live-trapping operations from late December to late February in the winter of 1964-65, and from mid-January to mid-March 1966. Efforts were made to trap as many of the Lamar (Crystal trap) and Mary Mountain (Nez Perce trap) animals as possible. More were trapped during the severe winter of 1964-65 than in 1966. Trapping success for calves of the previous spring was nearly 100%, as indicated by comparison of 1964-65 Mary Mountain early inter-aerial count records (72 calves) and Nez Perce trap records (77 calves). Most yearlings were probably trapped. Success (indicated by the temporary backtags and the number of retrapped animals) was high in the 2.5- and 3.5-year-old classes, but a few undoubtedly were not taken. Most adult females were trapped; trap success was lowest for males of age classes above 3.5 years, particularly in 1966. That year only 7% of all captures older than 3.5 years were males, compared with 37% in 1964-65 (Nez Perce trap). In addition to the influence of snow depth on trap success, animals trapped the previous year seemed harder to herd.
Age classes for all females trapped (Table 13) provided population sex-ratio information for animals less than 4.5 years of age. The records for all traps suggest that a slightly higher percentage of female calves survive their first year than do males, but the pooled data for yearlings and 2.5-year-olds suggest no further differential loss for this period. The variability, when individual trap records are compared, among the percentages of females in the 2.5- and 3.5-year-old classes may reflect the removal of calves in reductions of 1961-62 (Lamar) and 1962-63 (Mary Mountain). Although this may explain the very low percent of 3.5-year-old females taken at the Lamar trap, all trap records suggest that fewer females survive in this age group. Thus, female survival may be favored the first year, but male survival may be favored the next 3 years of life. Fuller (1960) also noted more males than females in the spikehorn, or 2.5- to 3.5-year-old group, but believed the identifications were biased. By 4.5 years of age and older, differential survival cannot be distinguished from differential trap success.
TABLE 13. Age classes of bison trapped, reductions of 1964-66.
| Lamar 1965-65 | Nez Perce 1964-65 | Nez Perce 1965-66 | Totals for both traps 1965-66 | |||||||||||||
| Females | Class | Females | Class | Females | Class | Females | Class | |||||||||
| Age Class | No. | % | No. | % Trap Total |
No. | % | No. | % Trap Total |
No. | % | No. | % Trap Total |
No. | % | No. | % Trap Total |
| Calf | 13 | 52 | 25 | 19 | 45 | 58 | 77 | 24 | 13 | 59 | 22 | 22 | 71 | 57 | 124 | 22 |
| Yearling | 10 | 53 | 19 | 14 | 26 | 50 | 52 | 16 | 11 | 44 | 25 | 25 | 47 | 51 | 96 | 17 |
| 2.5 | 4 | 40 | 10 | 6 | 17 | 68 | 25 | 8 | 3 | 27 | 11 | 11 | 24 | 52 | 46 | 8 |
| 3.5 | 3 | 21 | 14 | 11 | 8 | 42 | 19 | 6 | 5 | 45 | 11 | 11 | 16 | 37 | 44 | 8 |
| Adult | 34 | 53 | 64 | 49 | 97 | 63 | 154 | 47 | 28 | 93 | 30 | 30 | 159 | 64 | 248 | 44 |
| Totals | 64 | 49 | 132 | 100a | 193 | 59 | 327 | 100a | 60 | 60 | 99 | 100a | 317 | 57 | 558 | 100a |
aApparent errors in totals are due to rounding.
| ||||||||||||||||
Trap records, together with the early-winter aerial count of 480 animals, indicated that adult bulls outnumbered adult cows in the Mary Mountain population of 1964-65. The actual ratio could not be determined, but a minimum bull population of 150 was estimated from the number of trapped bulls removed (48), the scattered bulls in Hayden Valley which could not be driven toward the trap (53), and the number of bulls among the remaining 100 animals never trapped (50 bulls estimated). Because there were probably adult cows as well as some subadults of both sexes among the other 50 animals (of the 100 never trapped), bulls probably totaled less than 60% of the adult population. More bulls than cows could be expected in both the Lamar and Mary Mountain populations after the reductions of the early 1960s removed large numbers of adult cows. The normal proportion may be the reverse; adult cows may outnumber adult bulls. Fuller (1960) believed this to be the situation among the wild bison of Wood Buffalo National Park. Adult male bison may be "biologically expendable" according to Etkin (1964), who states that sexual dimorphism (termed aggressive potential in bison) interferes with male survival.
Age classes were calculated for the Mary Mountain population, using the Nez Perce trap records of 1964-65 and the prereduction count of 480 bison. Mary Mountain herd records were used, rather than pooled data, because trapping success was high (68%), the sample was largest, and trapping operations were preceded by a careful aerial census for the entire Firehole-Hayden Valley area. The calculations suggested a winter population consisting of: calves, 16%; yearlings, 11%; 2.5-year-olds, 6%; 3.5-year-olds, 5%; and adults, 62%. The extent to which these percentages differ from those of a Yellowstone population not influenced by reductions is not known.
Calf Percentages
Spring calf numbers, calculated as percentages of mixed herds (Table 14) from aerial surveys made in late May and early June 1964 through 1968 after most calves were born, showed considerable variation because of reduction programs. The lowest percentages recorded during the study for Lamar, Pelican, and Mary Mountain were 7, 14, and 7%, respectively, during the spring of 1965 after reductions in all three areas removed large numbers of cows. Thereafter, all percentages increased. By 1968 the Pelican population, where the fewest cows were removed showed what may be a nearly "normal" (uninfluenced by reductions) 18% calves. The higher percentages in Lamar (27%) and Mary Mountain (25%) for 1968 probably reflect the higher pregnancy rate of an increased proportion of younger females. Although the number of animals sampled was not large, the 1968 samples represented 95, 75, and 40% of the Pelican, Lamar, and Mary Mountain mixed herd groups.
TABLE 14. Calf percentages of mixed herd numbers in spring.
| Year | Lamar | Pelican | Mary Mountain | All herds | ||||
| No. sampleda | % Calves |
No. sampleda | % Calves |
No. sampleda | % Calves |
No. sampleda | % Calves | |
| 1964 | 43 | 19 | - | - | - | - | 43 | 19 |
| 1965b | 43 | 7 | 69 | 14 | 67 | 7 | 179 | 10 |
| 1966c | 40 est. | 15 est. | - | - | 109 | 12 | 149 | 13 |
| 1967 | 31 | 19 | 133 | 15 | 177 | 24 | 341 | 20 |
| 1968 | 34 | 27 | 111 | 18 | 48 | 25 | 262 | 19 |
aNumber sampled does not correspond with the actual population.
bReductions in all areas. cReduction in Mary Mountain herd. | ||||||||
Pooled percentages for the three herds in 1967 (20%) and 1968 (19%) suggest a leveling in calf production for the population as a whole. If so, then the usual proportion of newly born calves in mixed herds of Yellowstone bison may be 18-20%. Additional data, free from possible reduction influences, are needed to verify this.
Spring calves as a percentage of the total population (mixed herd groups, bull groups, and scattered animals) were calculated only in 1967 from a very accurate (397 count, 400 estimate) census of the entire park just before calving season. Calves formed only 11% of the 1967 spring total population, compared with 20% of the mixed herd groups. Although, as previously discussed, 20% calves in mixed herd groups may be nearly normal, 11% calves among the total population may be low. Reductions have removed a larger proportion of mixed herd animals than of bull groups and scattered animals. As a result, the total of mixed herd groups may form a smaller part of the 1967 population than it would of a population uninfluenced by recent reductions.
Mortality
Calf mortality
The expected number of calves from the 28 females of bleeding age released from the traps during the reduction of 1964-65, using the observed pregnancy rate of 52%, was 15. The actual number seen in those groups in the spring of 1965 was eight. The difference probably represented loss of embryos or neonates. A realistic percentage of such loss for the entire population could not be judged from the limited information.
Field observations of the relatively small bison population of the study period suggested that little additional calf mortality took place before late the following winter (March and early April). Throughout 1966 and 1967, during and after calving season in Lamar and Pelican, periodic ground and aerial checks were made. The agreement on total number of calves seen was quite consistent. Size and color of calves helped to confirm that loss, balanced by addition of some late calves, was not occurring. Calf percentages of herd numbers throughout the year (Table 15) were inconclusive, showing increases in 1965 and 1966, and slight decreases in 1964, 1967, and 1968. Size of some of the samples, skill of the observers, and variable conditions of weather and animal, visibility may have affected the quality of the counts. The percentages did suggest that there was no appreciable calf loss. Trap records for 1964-65 from Nez Perce showed a ratio of 61 calves per 100 cows (2.5 years of age and older) in winter, although among cows removed from the same population that winter the pregnancy rate was 51%. The sampling may have been biased, the previous year's pregnancy rate may have been higher (unlikely because of a large reduction in 1962-63), or mortality may have occurred in older age classes, but calf mortality was not indicated. In contrast, Fuller (1962) found a steady decline in calf percentages from a peak of 23% of herd numbers in July to less than 10% by the end of the calendar year.
TABLE 15. Calf percentages of pooled mixed herd numbers throughout the year.
| Spring | Summer | Fall | Winter | |||||
| Pop. yearb |
No. sampledb | % Calves |
No. sampledb | % Calves |
No. sampledb | % Calves |
No. sampledb | % Calves |
| 1963 | - | - | 908 | 17 | - | - | - | - |
| 1964 | 43 | 19 | 660 | 18 | 427 | 17 | - | - |
| 1965 | 179 | 10 | 591 | 13 | 242 | 17 | - | - |
| 1966 | 149 | 13 | 1533 | 19 | 121 | 20 | 165 | 21 |
| 1967 | 341 | 20 | 1724 | 19 | 78 | 18 | 40 | 18 |
| 1968 | 262 | 19 | 1003 | 18 | - | - | - | - |
aPopulation year includes spring, summer, and fall of the year shown,
and winter of the following calendar year.
bNumber sampled does not correspond with the actual population. | ||||||||
Subadult mortality
Trap records (Table 13) suggested considerable mortality among subadults. The records show that a progressive decline in numbers occurred between calf and yearling, and between yearling and 2.5-year-old age classes. One exception, an increase in percentage of yearlings compared with calves at the Nez Perce trap in 1965-66, may have been caused by a shift of a mixed herd group from one winter range to another. Previous reductions (1961-62 in Lamar; 1962-63 at Nez Perce) may have affected the size of some age classes, but do not explain the similar pattern of percentages among subadult age classes recorded at both traps in 1964-65.
The calculated age classes of the 1964-65 Mary Mountain winter population (16% calves, 11% yearlings, 6% 2.5-year-olds, 67% older) suggested that half the calves which survive into their first winter die before 2.5 years of age. Calf numbers indicated a potential 16% increase. Realized percentages for yearlings and 2.5-year-olds were 13% (11/84) and 8% (6/73), respectively. These indicate a 19% loss between the first and second winters, and a 31% loss between the second and third winters. This mortality most likely occurs in late winter (March and early April) at the end of the first and second years of life, rather than early in winter during the second and third years of life (see Other Mortality).
Variation in subadult mortality rates probably occurs, but cannot be judged from the limited data. Previous reductions, as discussed above, probably had less influence on subadult mortality than did environmental factors. Because this population had not been subject to an unusually severe winter for several years, these percentages may approximate usual mortality among these age groups.
Other mortality
Natural mortality recorded during the study period totaled 49 animals. All but one occurred during the winter after considerable snow had accumulated, usually after January. The number of recorded deaths were 38, 4, 3, and 3 for the winters of 1964-65 (a severe winter, 1965-66, 1966-67, and 1967-68, respectively. With the exception of a small mixed herd group (18 animals), all dead animals were found scattered on the winter ranges. Of the other 30 recorded deaths, there were 22 aged bulls, 2 cows, 3 yearlings, 1 calf, and 2 unknown. More bulls may have been found because of larger carcass size. Reductions, by removal of mixed herd animals, may have substituted for proportionately more natural mortality among cows and younger animals than among bulls.
Natural mortality recorded annually over mild and average winters of the study period averaged less than 1% of the winter herd numbers. One or two animals, usually bulls, were the most found dead in any one of the main wintering valleys. This was also the usual number found before the study period, according to earlier ranger reports, although population numbers varied considerably (Appendix IV). The amount of natural mortality associated with a severe winter such as 1964-65 was obscured by concurrent reduction programs, but the number of dead animals found indicated greatly increased mortality.
Population Trends
Figure 24 shows population trends based on actual counts (post-reduction, precalving) from 1936 through 1968 for the entire park, as well as for the wintering subunits of Lamar, Pelican, and Mary Mountain. Known losses, mainly reductions, are also shown. The reductions, held only in Lamar prior to 1954, accounted for the general downward trend in that area, although at the same time the population as a whole increased, influenced mainly by the steady increase of the Mary Mountain herd as it repopulated an area uninhabited by bison for more than 40 years. The peak population in the latter herd and for the total bison population in 1954 apparently coincided with increased use of the Madison Plateau-Pitchstone Plateau summer range, as mixed herd groups were small and infrequently reported from this area for nearly 15 years after the 1936 reintroduction on the Firehole. During the early 1950s, ranger reports indicate yearly use by mixed herds, sometimes large in size. In June 1952, 124 adults and 11 calves were seen.
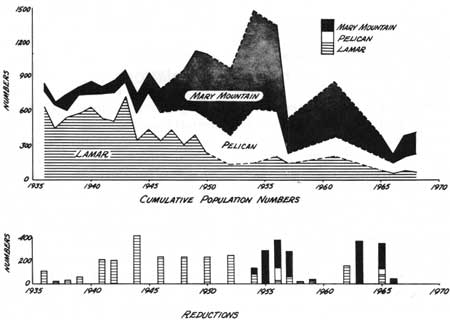
|
| Fig. 24. Bison population trends from population counts, 1936-68. |
Since the 1954 peak, all population segments have decreased. The 1955 reduction in the Mary Mountain area may have started the downward trend shown between the counts of 1954 and 1956. The marked decrease in population numbers in the Mary Mountain and Pelican herds, and the smaller decrease in Lamar cannot be entirely due to the reductions of 1956; there was none in Pelican. The decrease coincided with the severe winter of 1955-56, when snows were exceptionally heavy in January, and severe cold was prolonged in February. A large winter loss that year was indicated by sightings of 20-30 dead animals in Pelican during an aerial census of late January 1956 (Jim Stradley 1968 pers. comm.). The following June, 32 dead animals were counted in the main valley (Larry Brown 1969 pers. comm.). Reduction only added impetus, apparently, to a natural occurrence.
Immediate increase in the Lamar numbers followed reductions when population levels remained well over 200 animals, before 1952, by which time the last vestiges of the Buffalo Ranch operation (feeding of hay) ceased. Since then, Lamar has shown only very slow increase, although reductions have been few and comparatively small. Reported natural mortality (an occasional animal) indicated that no more bison than usual died most years. Slight increases which showed in counts of 1956 and 1961 were followed by reductions each time. At least 50% of the 1962 reduction consisted of females (yearlings and older). After the 1965 reduction, only nine females (yearlings and older) and 10 female calves were released. The temporary effect of even small reductions on low population levels was apparent, but some increase (with recruitment of more breeding females into the population) expected by 1968 did not occur. This suggests that some factor, other than reductions, may have retarded increase in this population at levels below 200 animals. The factor may be related to the change to a truly wild population, subject to natural environmental influences.
The Pelican population increased from near extinction about 1900 to an estimated more than 100 animals in the early 1920s. It was subject to reductions only twice, in 1956 and again in 1965. From the early 1920s through 1941, it fluctuated between counts of 100 and 200; for the next 10 years fluctuations were generally between 200 and 300 animals. After the peak count of 461 in 1954, followed by the drop to fewer than 100 in 1957, the population again remained between 100 and 200 animals. The lack of increase over fairly long periods of time suggests some balance exists with environmental factors, in which factors favoring increase are usually offset by mortality factors such as winterkill. The 1954 peak may have occurred with exceptionally favorable influences, which existed only briefly.
The park-wide population counts do not suggest that periodic emigration from Pelican to Lamar or Hayden Valley could account for the long-term stability, although apparently temporary population shifts have occurred (see Mixing of Population Segments). No herd groups were reported in Hayden Valley after 1895 until animals from Lamar were trucked there in 1936. Ranger reports made prior to the study period indicate that reproduction by herd groups in Lamar and Hayden Valley probably accounted for the usual population increases in those areas.
Population estimates for the entire park of 400 and 440 animals in 1967 and 1968, respectively, showed a 10% increment for that period. Reduction effects and insufficient population data precluded calculation of annual increment over a longer period. Between 1965 and 1968, when the study ended, the bison population for the park as a whole showed only slight increase. Three years had elapsed since a major reduction, and 2 years since a small reduction. The apparent increase in the Mary Mountain herd (Fig. 24) between 1966 and 1967 resulted from use of a low actual count in 1966 rather than the more accurate estimate. In all bison population segments, there is the suggestion of some factor or combination of factors retarding increase, aside from the influence of reductions.
Environmental conditions have continued to exert major influence on the bison populations in conjunction with the activities of man. Population trends (since the near extinction of the original wild population about 1902) apparently reflect, initially, reestablishment of populations on historic ranges which had been uninhabited by any bison for some years (Lamar, Mary Mountain), or where the numbers were very low (Pelican). Increase was rapid under conditions of semidomestication (Lamar), and when favorable environmental conditions combined with availability of an extensive, uninhabited area (Mary Mountain). Later increases were apparently offset by reductions (Lamar), or by reductions in combination with periodically severe environmental conditions (Pelican, Mary Mountain). At lower population numbers, less severe environmental conditions appear adequate to have offset increases and to have resulted in periods of near stability or very slow increase in all population segments.
Presumably, the original wild bison population which inhabited the Yellowstone area throughout the year lived in some balance with the prevailing environmental conditions of that time. Fluctuations in numbers undoubtedly occurred as favorable conditions alternated with periodically severe winters. The present environment of the bison ranges of the park appears little altered by man's influence (with the possible exception of Lamar Valley), although the numbers of some of the predatory and grazing mammals have decreased. The bison population may be expected to increase in the future (since population numbers of the study period are the lowest in many years after the reductions of the 1960s), but may then establish a state of equilibrium with natural environmental conditions.
| <<< Previous | <<< Contents >>> | Next >>> |
chap5.htm
Last Updated: 24-Jan-2005