|
YELLOWSTONE
The Bison of Yellowstone National Park NPS Scientific Monograph No. 1 |

|
CHAPTER 7:
HABITS
Calving
COMPARISON OF RECORDS kept during the study period with earlier information suggested that the onset of the calving season is later than formerly. From 1937 to about 1950, the first calves were regularly reported in Lamar before mid-April. Since 1950, first calves have all been reported later. McHugh (1958) reported that calving season extended from 15 April to 31 May, with more births the first 2 weeks of May. During the study period most births occurred during the first half of May, and calving season was over by the end of May. There were always a few late births; occasionally a new calf was seen in late summer. (Subsequent to this study, calving season has extended into June as the population has increased.)
Fetal size information compiled before the study period also suggested that a large part of the calving extended over a longer period of time. Rush (1932a) reported only that there was considerable variation. E. R. Quortrup's unpublished data (Skinner 1941) indicated a size range from a 2-inch embryo to a 23-pound, 22-inch fetus; 80% of them weighed between 5 and 20 pounds when examined in late January (Appendix V).
In all Yellowstone herds during the study period, most calves were born at about the same time (early May). Observations of the earliest calves were rarely possible except in Lamar (because of limited access before 1 May), but the few records made by rangers for the Firehole suggested that first calves are usually born there perhaps a week later than in Lamar.
Calves were born on the wintering areas (Fig. 28), except for the occasional late one. In Lamar during this study, most calving took place in the lower Lamar Valley (west of Lamar Canyon in the area called the Horseshoe or Little America). Ranger reports predating the study indicated that this same area was the usual place of calving. In Pelican, most calving occurred in the eastern half of the main valley from Astringent Creek to Raven Creek. Mary Mountain animals calved in both Hayden and Firehole valleys. On the Firehole, most calves were born in the Lower Geyser Basin areas of Sentinel Creek, Fairy Meadows, Pocket Basin, lower Nez Perce Creek, and the Firehole Loop. Most Hayden Valley calving occurred in the western half of the big valley, near Alum Creek.

|
| Fig. 28. Calves (a few days old) born about 1, May 1970, when snow was on the ground later than usual. This cow was observed deliberately pushing these calves onto their feet—an action not seen other years. Photo by Mike Sample. |
Rut
Most breeding activity occurred from mid-July to mid-August. Field observations throughout the study indicated that every year, for at least a part of that time, most of the herd groups which wintered in Lamar and Pelican intermixed in the Upper Lamar area. At the start of the rut in 1967, all of the animals were at the crest of the Absaroka Mountains on the east boundary, between the Hoodoos and Canoe Lake. In other years the groups were more scattered, some on the boundary and the adjacent ridges between Cache and Miller creeks, and some on Saddle Mountain. By the last week of July or the first week of August, some of these animals crossed the Lamar River to the Mirror Plateau, where breeding activity occurred from the meadows south of Amethyst Mountain south to the meadows at the headwaters of Pelican-Timothy-Raven creeks. Most of the breeding activity in the Mary Mountain herd occurred in the southwest corner of Hayden Valley and adjacent forest and meadow areas. In all of these areas there were small uprooted conifers and many rubbed trees. Small wallow-sized patches of grass (Fig. 29), apparently uprooted by morning but not much used for wallowing, were another common sign of rutting activity.

|
| Fig. 29. Grassy spot uprooted by bison horning during the rut. |
The period of breeding activity observed during this study (1963-68) was shorter than that given by McHugh (1958). He observed in the much larger Hayden Valley population of that time that the season extended from 15 June to 30 September, with less activity during the first and last 2 weeks.
Movements
Migratory movements
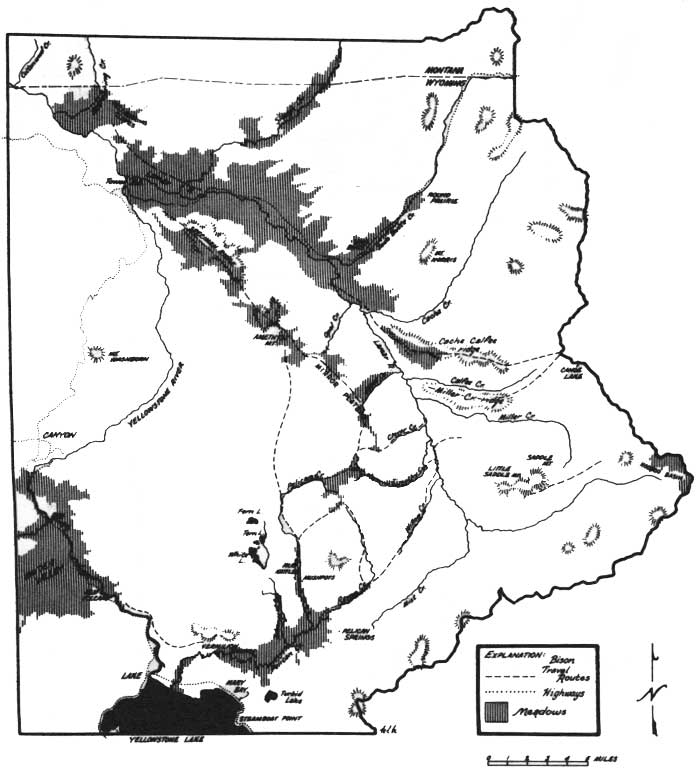
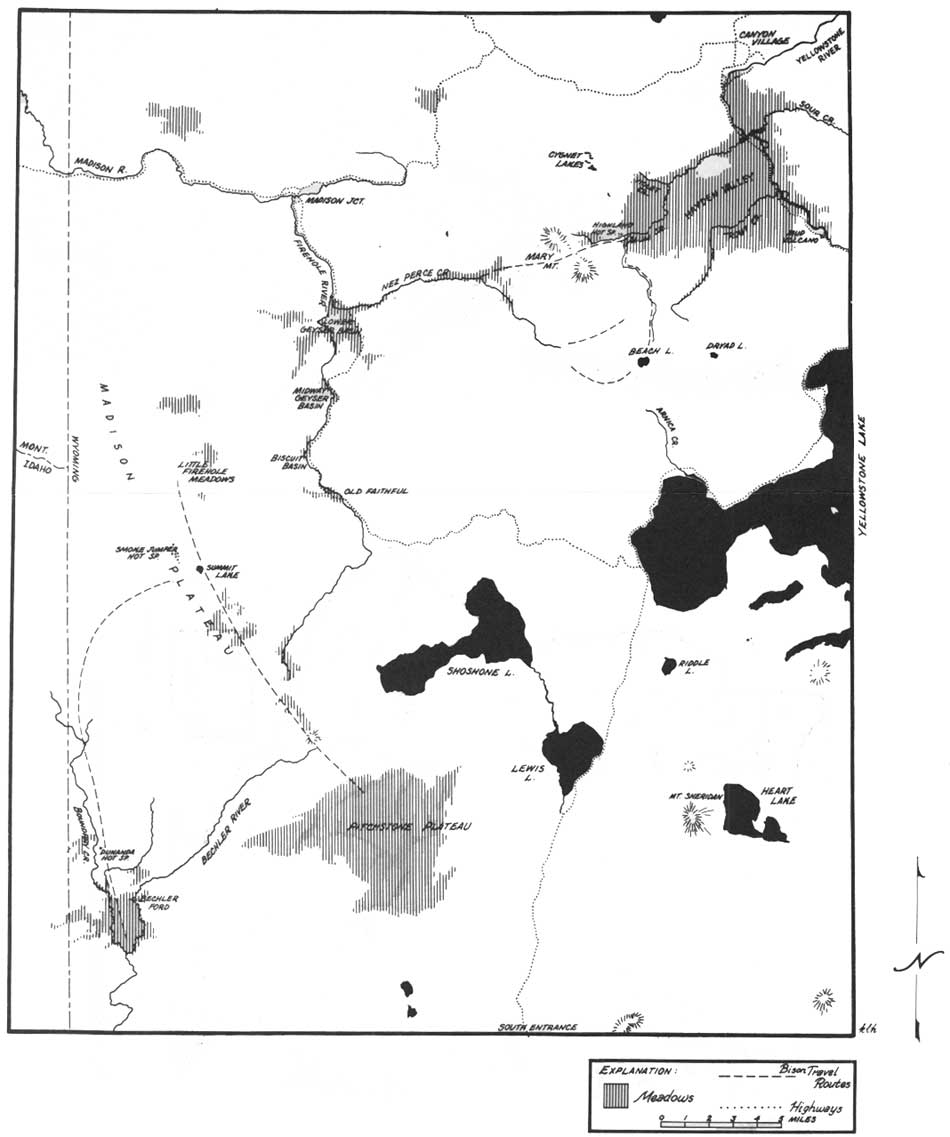
Most of the bison in Yellowstone are migratory, moving in spring from the lower wintering valleys to higher summer ranges, and reversing this altitudinal migration in the fall. The migratory pattern was most apparent in the mixed herd groups which wintered in the Lamar, Firehole, and Pelican valleys. Although all groups of a particular wintering area did not move at once in spring, they all moved within a few days' time. Records kept during the study showed that mixed herd groups wintering in Lamar moved 12-15 miles to higher areas of Mount Norris and the Cache-Calfee Ridge during the first week of June, 3 of 5 years, and by the last week of May twice. Firehole animals moved to Hayden Valley or the Little Firehole Meadows area of the Madison Plateau during the first week of June, 3 of 4 springs. Wintering groups began to move from lower to upper Pelican Valley at the same time. Some years, further movement across the Mirror Plateau to the Upper Lamar area was delayed until mid-June. Observations suggested that nearly every year the entire Pelican population, except some bulls, crossed to the Upper Lamar, returning to the Mirror later in the summer. Most movements took place along definite travel routes, as shown in Figs. 30 and 31.
|
| Fig. 30. Map of the Lamar-Pelican area of Yellowstone National Park. (click on image for an enlargement in a new window) |
|
| Fig. 31. Map of Hayden Valley-Firehole-Madison Plateau area of Yellowstone National Park. (click on image for an enlargement in a new window) |
The bulls that wintered as solitary animals, or in bull groups, left the wintering areas gradually during the spring. The last of them disappeared several weeks after the mixed herd groups moved. A few bulls were usually seen from the main roads in the Firehole, Hayden Valley, at Mary Bay, and in Lamar Valley almost to the end of June. The bulls followed not only the routes used by the mixed groups but moved upcountry along the various stream drainages to scatter widely on the summer ranges.
The cause of the spring movement was not clear. In 1966, an earlier spring than others of the study, the mixed herd groups moved from all three valleys by 24 May. In 1967, the Pelican animals were observed sequentially from the air. They moved, in 2 days, the 15 miles from the new spring growth of the upper Pelican Valley, across high meadows, which were mostly covered with snow or standing water, to the Lamar River. By the third day they were on the greening slopes of Little Saddle Mountain. They did not follow any discernible vegetation or snow melt sequence. Spring weather patterns and temperatures presumably influenced the migration. Dave Pierson (1968 pers. comm.), after years of observation, believed that warm spring days were the main factor.
In the late fall the movement of mixed herd groups was reversed, but occurred less abruptly than in the spring. Lamar animals appeared in the main valley (to stay) by late November. Some groups were reported in the Lower Geyser Basin about the middle of September most years, although the majority arrived in late October or early November. Pelican animals moved down sometime in the first 2 weeks of November, although groups commonly appeared earlier for a few days at a time when fall snowstorms occurred. This intermittent shifting from summer to winter ranges during stormy periods in the fall was also seen in Lamar.
Bulls commonly began to appear on winter ranges before herd groups were reported. The movement was gradual, continuing for some time after herd groups moved in for the winter. Most winters, a few solitary bulls stayed at sites high on the summer range, such as the heads of Opal and Flint creeks.
Cold temperatures alone did not cause shifts from summer to winter ranges; immediately after a fall storm, in temperatures often near 0°F, herd groups which had moved down returned to high elevations. Snow depth and exposure to storm conditions at the higher elevations apparently caused both the temporary and the final fall movements to winter ranges.
Summer range movements
During the summer, there were extensive shifts of all mixed herd groups on the various ranges. Animals which moved west onto the Madison Plateau from the Firehole, although few in number during the study, commonly used the Buffalo and Little Firehole meadows in early summer. From there, they moved south along the Continental Divide onto upper Boundary Creek and the western end of the Pitchstone Plateau, remaining in these latter areas un til the fall move to winter ranges.
Hayden Valley herd groups commonly stayed within the main valley much of the summer (Fig. 32). Other years, by July, they ranged the small meadows and parks scattered in the extensive lodgepole forest (Fig. 33) south to Dryad and Beach lakes, west to the upper reaches of Spruce Creek, and across Mary Mountain to upper Nez Perce Creek. Bulls from Hayden Valley occasionally moved south down Arnica Creek to West Thumb and on toward South Entrance.

|
| Fig. 32. Bison herd groups on Alum Creek in the western part of Hayden Valley. |

|
| Fig. 33. Bison on the strip of sedge meadows along upper Alum Creek south of Hayden Valley. |
Pelican and Lamar wintering herds were seen in early summer on the lower ridges of the Upper Lamar (Mount Norris, the western ends of the Cache-Calfee and Miller Creek ridges, and Little Saddle Mountain). By the end of June to mid-July, they moved higher, to the east boundary (Fig. 34), ranging from the Hoodoos to Canoe Lake and Saddle Mountain. By late July to early August, groups usually moved west down the ridges, and some crossed the Lamar River to the Mirror Plateau, where they ranged the northeast rim from Flint creek to upper Raven Creek until the shift to winter range. At higher population numbers than those occurring during this study, groups also commonly moved the length of Specimen Ridge on the north edge of the Mirror Plateau (Dave Pierson 1968 pers. comm.).

|
| Fig. 34. A herd group on the east boundary near Canoe Lake in early August. Photo by John Good, Yellowstone National Park. |
Apparent influences for movement of groups over large areas during summer were the rut, seasonal vegetation changes (see Use of Forage Areas), and, most obviously, biting insect populations. Small, gray, biting flies were particularly noticed on the Mirror and the Upper Lamar in 1965. In 1967, the flies were numerous in those areas, and in Hayden Valley also. From mid-June to mid-August 1965, neither aerial flights nor ground checks showed mixed herds on the Mirror Plateau, although in 1966 groups were seen there by 24 July. From mid-June to early August 1967, herd groups were observed only once in Hayden Valley, but were seen, from the air, to the south. Every year herd groups were seldom observed in Hayden Valley during much of July when biting insects were at their worst.
Burger (1967a) compared biting insect populations and animal distribution in 1966 and 1967. He noted that the distribution and abundance of species of Symphoromyia, a small gray biting fly, particularly at elevations below 8500 feet, was much greater in 1967. He suggested that this influenced the concentration of the entire Upper Lamar population of bison groups high on the east boundary, at elevations of 9500-10,000 feet, for more than 2 weeks in late July and early August 1967. Other summers, although the same boundary area was used by bison, they did not remain as long and were more broadly distributed.
Apparently, the insect populations influenced bison distribution and concentration more strongly than did breeding activity. In 1966, with fewer biting flies, mixed herd, groups split and moved from the Upper Lamar to the Mirror while the rut was at peak. In 1967, a similar move did not occur until both the insect populations and the rutting activity began to diminish.
The spring and fall migratory habits of the Yellowstone bison did not prevent animals from occasionally visiting areas of winter range during the summer. Observations made during the study period and earlier ranger reports indicated that scattered bulls sometimes appeared on all winter ranges. When population levels were higher, mixed herd groups also occasionally visited winter ranges for brief periods, according to ranger reports. Such groups were sometimes seen in the main Lamar Valley in mid-July, or more commonly, in August. Mid-summer movements of groups rarely occurred to areas near roads in the Firehole. No groups were reported near the main road through Hayden Valley in mid-summer, nor in the main Pelican Valley. No reason for these mid-summer movements was apparent.
Winter range movements
During the winter, all bison (mixed herd groups, bull groups, and solitary bulls) moved shorter distances and less frequently than in the summer. Movement was usually confined to the limits of the wintering valleys (see Numbers and Distribution), with egress difficult if not impossible after considerable snow had fallen. Mixed herd groups moved most frequently and for greater distances, but sometimes stayed in one locality for days or even weeks. Bull groups occupied smaller areas for longer periods. Solitary bulls sometimes spent the entire winter at one site. Presumably, the amount of movement was usually related to group food requirements. Winter conditions hindered but did not prevent occasional extensive movements by groups and single bulls between main wintering valleys. Reasons for such moves were not apparent, but forage requirements did not appear to be a factor. No movements to areas of high summer range during the winter are known; the solitary bulls wintering in these areas arrived in fall.
Mixed herd groups moved most frequently between Hayden Valley and the Firehole. In January 1965, after attempts to drive animals from Hayden Valley to the Nez Perce Creek trap with helicopters failed because of the above-average snow depths, most of the herd animals crossed of their own volition a few days later. The following winter, during the 1965-66 reduction, neckbanded animals released from the trap were seen a few days later in Hayden Valley. Before the present study, in March 1945, 54 bison traveled 6 miles across Mary Mountain in 42 inches of snow, according to McHugh (1958). The frequency of such crossings, uninfluenced by reduction operations, was unknown.
Mid-winter movements between Lamar and Pelican valleys apparently involved only bulls. Sometime between early December 1965 and mid-January 1966, a marked young bull crossed from Lamar, presumably by way of the Mist Creek divide. Rangers reported movement on this same route by bulls in February 1932 and January 1949. Jim Stradley (1968 pers. Comm.) saw a few mixed herd groups crossing from Pelican into Lamar in early winter about 1955-56.
Movements between Pelican and Hayden valleys apparently occurred in the fall and early winter, but sightings were rare. The trails of solitary bulls were followed from the air in January 1968. Mixed herd groups were reported only once during the study period east of the Yellowstone River near the Mud Volcano, but earlier reports indicated an occasional group in late fall. Jim Stradley (1968 pers. Comm.) observed from the air about two dozen cows and calves moving between the two areas about 1956-57.
Movements beyond park boundaries
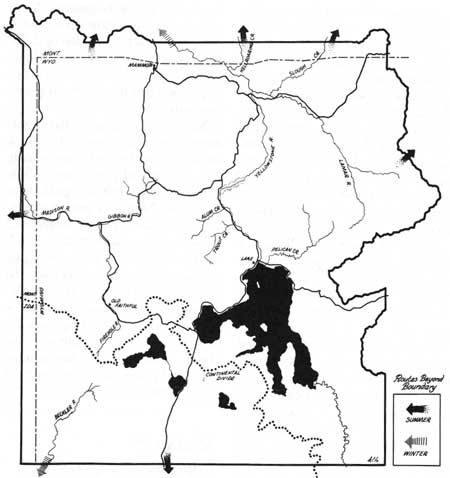
Since the early years of the park, few bison have moved beyond the boundaries (Appendix VI). Figure 35 shows the locations of reported movements, which usually involved one or more bulls. Most years some bulls summered beyond the north boundary. Since 1951, a few bulls have been reported almost every year beyond the southwest boundary in Idaho. These movements have occurred regardless of fluctuations in population numbers.
|
| Fig. 35. Map showing known locations of bison movements across the boundaries of Yellowstone National Park. |
Mixed herd groups have only been reported beyond the north boundary of the park. Late during the winters of 1943 and 1948, groups briefly left the park near Gardiner, Montana. Severe weather conditions partly influenced these moves. In summer 1943, nearly 150 bison were seen just outside the northeast corner. In the fall of 1947, 31 were 8 miles north of the boundary on Elk Creek in the Hellroaring area (R. Murphy 1970 pers. comm.). All these mixed-group movements occurred when larger numbers of bison wintered in Lamar.
Mixing of Population
Segments
Census records, aerial observations, and the limited records of neckbanded bison (Fig. 36) indicated that movements of population groups affected numbers inhabiting the winter ranges from one year to the next (population shifts), and resulted in interbreeding between populations of wintering areas during the summer. Movement occurred during the fall and winter between the valleys of the Firehole, Hayden, Pelican, and Lamar, as previously discussed. Observations confirmed the assumption made by, park personnel some years ago that the extent and frequency of mixing between the Firehole and Hayden Valley animals during most of the year, including the breeding season, justified considering them as one population, called the Mary Mountain herd, distinct but not isolated from the other two herds.

|
| Fig. 36. Neckbanded bison in a group at Pocket Basin on the Firehole. Photo by John Douglass, Yellowstone National Park. |
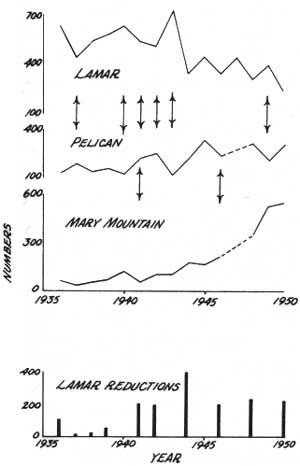
Shifts of other population groups were indicated by some fluctuations in actual winter counts from 1935 to 1950 (Fig. 37). Decreases in Lamar winter numbers coincided with increases in Pelican in 1937, 1941, and 1942; decreases in Pelican winter numbers coincided with increases in Lamar in 1940, 1943, and 1949. Reductions, made only in Lamar during this period, probably added to the amount of decrease there in 1941 and 1942, but could not have influenced the 1937 decrease nor any Pelican decreases. Bias in Pelican population counts made by rangers on winter ski trips might account for some variation, but not the coincidence of changes in population numbers in the two valleys. Mixed herd groups which moved between Lamar and Pelican in late fall and early winter probably remained on the winter range that was in use when weather conditions worsened.
|
| Fig. 37. Population fluctuations in wintering areas caused by shifts of mixed herd groups. |
Shifts between Pelican and Hayden Valley were infrequent and less obvious. Such shifts did occur in 1941 and 1946 (Fig. 37). Other shifts have occurred since, but the less frequent population counts made after 1950 gave no indication. During the study period, aerial observations and reduction records suggested two shifts occurred. Mary Mountain population counts after the 1964-65 reduction totaled fewer than the expected number of animals, based on numbers seen before trapping began (480), numbers removed (238), and possible natural mortality. A move of animals to Pelican after the early December count would explain the discrepancy. The next winter, in the reduction of 1965-66, 47% of the animals (yearlings and older) trapped at Nez Perce had no metal eartags, showing they had not been trapped the previous year. A movement into Hayden Valley from Pelican was the probable explanation.
Apparently, the shifts indicated in Fig. 37 were temporary, usually for the one winter. The presence of a mixed herd group (34 animals) wintering in Bechler Meadows in 1962-63 also indicated this. No herd group was reported there in 1961-62 or 1963-64. Over a span of time, the wintering numbers of the Mary Mountain, Pelican, and Lamar population segments would be unaffected by such shifts.
Observations of both total numbers of animals and individuals neckbanded in Lamar showed that, during the breeding season, intermingling of the Lamar and Pelican wintering groups was extensive most years. In 1967, all of them mixed together in several large, flexible groups at the crest of the Absarokas for more than 2 weeks. The amount of interbreeding which presumably would occur in such circumstances suggested that these bison also be considered one population segment. However, because winter season movements were so few, the separate designations were retained.
Separation was most complete between the Mary Mountain population and the Pelican-Lamar animals. The infrequent movements between Pelican and Hayden Valley at any time, coupled with the separation of the population segments during breeding season, resulted in almost no intermixing.
Historically, when there were large summering populations (parts of which wintered beyond the park area) north of the Lamar and west of the Firehole, interbreeding between the wintering populations of the Lamar and Pelican valleys, and of the Firehole and Hayden valleys, was probably less extensive because of geographic separation of some groups during the summer. Then as now, however, the groups which shared summer ranges undoubtedly interbred. Before intensive poaching greatly reduced numbers and probably altered or eliminated habitual patterns of movement, there may have been more contact between Pelican and Hayden Valley animals, but the amount of intermingling and interbreeding was probably least, even then, of the population segments.
Geographic separation during the breeding period through seasonal migration to higher summer ranges, seen in the present park bison on a local scale, could explain the existence of two recognized subspecies of bison in historic times whose distribution presumably overlapped in the Rocky Mountains (Skinner and Kaisen 1947). Possibly the mountain and plains types jointly used some wintering places along the east side of the Rockies, but breeding isolation kept the populations distinct.
The apparent temporary nature of group population shifts between main areas of winter range, discussed previously, and neckband records both suggest that animals have affinities for particular winter ranges. Neckband records for the Mary Mountain herd showed that some individuals preferred one winter valley to the other, regardless of movement between the two valleys at all seasons. The entire population (except for the many scattered bulls) had crossed to the Firehole side during the winter of 1964-65 when helicopters drove them toward the trap on Nez Perce Creek. Most summered (1965) in Hayden Valley. Those in Hayden Valley when trapping operations again began during the winter of 1965-66 were driven across to the Firehole side to the trap on Nez Perce Creek. Aerial observations after the trapping showed that several bison, neckbanded and released, recrossed the divide to Hayden Valley immediately. During the winter of 1966-67, when there was no trapping, five marked animals (neckbanded in 1965-66) wintered in Hayden Valley: an adult cow, a 4-year-old cow, a 2-year-old cow, and two young bulls. Three marked adult cows wintered on the Firehole in both 1966-67 and 1967-68 although they were identified in Hayden Valley during summers. Less complete records on other marked cows also suggest affinity for a particular winter range although most mixed herd groups summered in Hayden Valley.
Records for the animals marked in Lamar further suggest that the tendency to winter habitually in the same valleys was stronger among mature females. Marked animals of all ages were seen on various parts of the summer range, including Raven Creek on the Mirror Plateau (close to Pelican Valley), but only three immature bulls and two immature females spent at least one winter in Pelican Valley. Cows marked as adults (five marked) were identified in winter only in the Lamar Valley, two of them for three successive winters.
A tendency for animals, particularly mature females, to prefer a given winter range might explain the lack of any lasting interchange or permanent shifts by groups from one winter range to another. The lack of emigration to new territory, such as from Pelican to Hayden Valley before the reintroduction of 1936, might also be explained. Possibly, except for young animals or old bulls, interchange between wintering valleys involved only groups. Although the basis of such groups is not known, observers generally agree that a group structure exists. Fuller (1960) believed there was a unit of some sort. McHugh (1958) believed the groups were flexible and depended little on blood relation. He did observe that cow (mixed herd) groups were usually led by mature females. These cows would then govern the movements made by the groups.
Bison have shown willingness to move considerable distances in rough topography or deep snow, according to the time of year, but they do not all intermix freely. Their movements follow repetitive patterns which indicate habit as well as physical need. The movements of both individuals and groups result in a varying amount of separation among population segments which winter in the valleys of the Firehole, Hayden, Pelican, and Lamar (Table 16). The Yellowstone bison of the present (and recent past) remain almost entirely within the park boundaries throughout the year. Those that range to the northeast (Lamar and Pelican) are almost separate from those designated the Mary Mountain herd. Affinities for given winter ranges are responsible for much of the separation, which is reinforced by migratory habits which maintain a breeding season isolation. Occasional movements by bulls and mixed herd groups prevent total separation of any one part of the population.
TABLE 16. Comparative summary of movements shown by the present wintering populations.
| Firehole-Hayden Valley Populations |
Pelican-Lamar Populations | |||
| Winter separation | Partial | Almost complete | Almost complete | |
| Group movement between valleys | Frequent - year around | Least frequent - fall and early wintera | Infrequent - fall and early winter | |
| Seasonal migrations | Less marked | Quite marked | ||
| Summer interbreeding | Complete | Complete | ||
| Amount of movement on summer range | Range less widely (except Madison Plateau) | Range widely | ||
| Amount of movement on winter range | Varies among herd groups, bull groups, and solitary bulls | Varies among herd groups, bull groups, and solitary bulls | ||
| Movement beyond boundaries | A few bulls, SW corner | A few bulls, N&E boundaries, summer; herd groups, N boundary, winter | ||
| Summary: | ||||
| Population separation | One population: Mary Mountain | Almost separate | One breeding population, with winter range designations as Lamar and Pelican | |
aApplies to relationship between Hayden and Pelican valleys.
| ||||
Food Habits
Results of analysis of 22 rumen samples, half of them collected in the winter and half throughout the rest of the year, are shown in Table 17. The 1-quart sample taken represented approximately 1% of the rumen-reticulum volume of 91 quarts averaged from eight adult males. Approximately two-thirds of the samples were taken from adult males. Because of some uncertainties in distinguishing between sedges and some grasses, doubtful material was placed in the grass category. Sedge may thus be underrepresented in the analysis.
TABLE 17. Food habits as indicated by rumen sample analysis.
| Species | Winter 11 samples | Spring 4 samples |
Fall 3 samples | Total 22 samples | ||||||
| Freq. % | Comp. % | Freq. % | Comp. % | Freq. % | Comp. % | Freq. % | Comp. % | Freq. % | Comp. % | |
| Grasses and grass-like plants | 100 | 99 | 100 | 96 | 100 | 91 | 100 | 99 | 100 | 96 |
| Sedge | 100 | 56 | 100 | 49 | 100 | 50 | 100 | 37 | 100 | 51 |
| Grasses | 100 | 34 | 100 | 46 | 100 | 32 | 100 | 30 | 100 | 35 |
| Wire rush | 100 | 9 | 100 | 1 | 100 | 8 | 100 | 32 | 91 | 10 |
| Spike-sedge | -- | -- | -- | -- | 50 | 1 | -- | -- | 9 | trace |
| Forbs | 45 | trace | 100 | 3 | 100 | 6 | 67 | trace | 68 | 2 |
| Phlox | 18 | trace | 75 | 2 | 25 | trace | -- | -- | 27 | 1 |
| Northwest cinquefoil | -- | -- | 25 | trace | 100 | 6 | -- | -- | 23 | 1 |
| Sulfur eriogonum | 9 | trace | 100 | trace | 75 | trace | 33 | trace | 36 | trace |
| Dandelion | 73 | trace | -- | -- | 75 | trace | 33 | trace | 32 | trace |
| Pussytoes | -- | -- | 75 | trace | -- | -- | -- | -- | 14 | trace |
| Groundsel | -- | -- | -- | -- | 75 | trace | -- | -- | 14 | trace |
| Shrubby cinquefoil | 18 | trace | -- | -- | -- | -- | -- | -- | 9 | trace |
| Clover | -- | -- | -- | -- | 50 | trace | -- | -- | 9 | trace |
| Onion | -- | -- | -- | -- | 25 | trace | -- | -- | 5 | trace |
| Blue-eyed Mary | -- | -- | -- | -- | -- | -- | 33 | trace | 5 | trace |
| Unidentified | -- | -- | 25 | trace | 75 | trace | 33 | trace | 23 | trace |
| Browse | 82 | 1 | 75 | trace | 50 | 2 | 67 | trace | 73 | 1 |
| Big sagebrush | 36 | 1 | 50 | trace | -- | -- | -- | -- | 27 | 1 |
| Red dogwood | 45 | trace | -- | -- | -- | -- | -- | -- | 23 | trace |
| Raspberry | 9 | trace | -- | -- | 25 | trace | -- | -- | 9 | trace |
| Dwarf huckleberry | -- | -- | -- | -- | 25 | 2 | -- | -- | 5 | trace |
| Serviceberry | -- | -- | -- | -- | -- | -- | 25 | trace | 5 | trace |
| Fringed sagebrush | 9 | trace | -- | -- | -- | -- | -- | -- | 5 | trace |
| Unidentified | 45 | trace | 25 | trace | -- | -- | 67 | trace | 36 | trace |
| Lodgepole pine | 18 | trace | 75 | trace | 50 | trace | 33 | trace | 36 | trace |
| Horsetail | 18 | trace | 25 | trace | 25 | trace | -- | -- | 18 | trace |
| Moss | 9 | trace | -- | -- | 25 | trace | 33 | trace | 14 | trace |
| Lichen | -- | -- | 25 | trace | -- | -- | -- | -- | 5 | trace |
| Unidentified | 36 | trace | 75 | trace | 50 | trace | 33 | trace | 46 | trace |
Trace indicates less than 1% composition.
Material is ranked according to composition by volume, in the total diet. Composition totals of less than 100% are the result of rounding to the nearest percent. | ||||||||||
Grasses and grasslike plants formed most of the diet throughout the year. Totals were 99, 96, 91, and 99% for winter, spring, summer, and fall, respectively. Sedge was the main source of forage at all seasons, averaging more than half the diet. Its value in winter (56%) reflects the animals' frequent concentration during this season on sites where nearly all the plant growth is sedge. Grass, second in quantity during all seasons except fall, averaged more than one-third of the diet for the entire year. Its highest value (46%) in spring probably reflects both greater palatability and increased availability of new spring growth. Wire rush was taken yearlong, but averaged approximately one-third of the fall diet.
Both forbs and browse were represented throughout the year in the bison diet, usually as trace items. Forbs (mostly phlox) averaged 3% of the spring diet; northwest cinquefoil averaged 6% of the summer diet. Although forbs did not total a large percentage of the forage utilized, they may be an important part of the diet nutritionally.
Browse was of least importance quantitatively, forming only 1% of the year's diet. Six species were identified—four as trace values. Dwarf huckleberry represented 2% of the summer diet but occurred in only one of the four samples. Its presence may be incidental to the summer use of forested areas where it is common on the forest floor. Other browse plants shown in the analysis may also be incidental to feeding at a particular site. Their possible importance nutritionally is unknown.
Forage composition data for the bison use areas represented by the food-habits analysis are limited. Descriptions of species composition for the Lamar Valley are contained in Rush (1932b). The Soil Conservation Service (1963) and Bergstrom (1964) provide some quantitative data. Descriptions of species composition and abundance at higher elevations of the Lamar area were made by Oldemeyer (1966) and Woolf (1968). Hayden Valley species are mentioned by Kittams (1949) and the Soil Conservation Service (1964). From these sources and information obtained during this study, a list of grasses and grasslike plants was compiled according to site (Appendix VII). Forage composition information for the thermal sites used by bison during critical winter periods (see Use of Thermal Areas) has not been determined.
Wet bottomlands, creek banks, and pond edges support dense sedge growth (mainly Carex aquatilis, beaked sedge, and Nebraska sedge). Slightly less wet sites support a greater variety of sedges, horsetail, wire rush, and grasses such as sweetgrass, timothy (particularly in Lamar), and tufted hairgrass.
Moist meadows of the wintering valleys have some of the same sedges (Carex acuta, C. platylepsis, C. raynoldsii) found in the less wet sites such as swales. Smallwing sedge was the most commonly observed additional species. Horsetail, rushes, and additional grasses, particularly bluegrass, are found. Giant wild rye is prominent in Lamar.
Moist and dry upland sites both support bluegrass species, Idaho fescue, and needlegrass. On the drier sites, junegrass and various wheatgrasses occur in most areas; the sedge Carex xerantica is common in Lamar.
At higher elevations on mountain herbland sites, sedges such as Carex raynoldsii and smallwing sedge are prominent. Rushes, bluegrasses, Idaho fescue, mountain brome, needlegrass, wheatgrass, alpine timothy, and tufted hairgrass are all common. Subalpine meadows have fewer sedges (mainly Carex raynoldsii and Hepburn sedge). Grasses are predominantly bluegrasses and tufted hairgrass.
| <<< Previous | <<< Contents >>> | Next >>> |
chap7.htm
Last Updated: 24-Jan-2005