|
NATIONAL PARK SERVICE
The Giant Sequoia of the Sierra Nevada |

|
CHAPTER 2:
THE TREE AS AN INDIVIDUAL
Description
Human propensity for superlatives has led to a rather one-sided view, as the giant sequoias described were mainly the larger specimens. Perusing the literature might lead us to suspect that young trees were scarce indeed, and seedlings nonexistent.
Seeing the trees themselves, we realize that descriptions are inadequate to do them justice. Records of the various measurements fail to convey the trees' massive beauty, and even pictures cannot reproduce their awesomeness.
Writers have long been inclined to compare the tree with other objects of known dimensions. The number of horses and riders that could stand in the hollowed cavern of the trunk, the arm lengths required to span its width, or the number of houses that one could build with its wood are familiar examples of such comparisons. Hinds (1893) declared that a mature sequoia tree is 3,125,000,000,000,000,000,000 times larger than a single bacterium—a comparison of questionable value, the author neglecting to say which sequoia was meant, and which species of bacterium.
As stated earlier, the erroneous and exaggerated sizes and ages of these trees have been refuted in recent years through the use of standardized measurement procedures and more sophisticated instruments. Despite this, standard procedures are not necessarily followed and reference points from which measurements were made are not necessarily given. The dimensions used in this account are official figures obtained from the National Park Service and the U.S. Forest Service.
Because the tree's appearance varies greatly with age, we will discuss each major stage of its life cycle.
Seedling stage
As germination progresses, the emerging sequoia seedling stands from three-quarters of an inch to an inch high by the time the seed coat is shed from its spreading cotyledons (Fig. 8). The number of cotyledons or seed leaves, varying from three to six, is usually four. They are bright green on the upper surface, reddish on the underside, and rather sharp-pointed. To the novice observer, they may closely resemble the seedlings of the white fir. The latter's seed leaves, however, are bluer in color, have blunter tips, and green undersides, all of which clearly distinguish them from the neighboring young sequoias.

|
| Fig. 8. Two-week-old sequoia seedling. At this age, the typical seedling is about 0.75 inch tall, and bears four cotyledons of seed leaves. Depending upon soil moisture conditions, secondary leaves will make their appearance within 2.5 weeks. |
Where site conditions approximate the optimum and the seedlings survive, the linear secondary leaves normally begin to appear within a matter of weeks as a rosette in the center of the developing crown. Depending upon soil moisture conditions, the cotyledons wither and fall away within a few to several weeks. Branching begins by late summer and the more vigorous specimens may have as many as six branches by autumn when the plant is perhaps 3 or 4 inches tall. By this time, the leaves of the newer foliage begin to resemble the awl-shape of those on the adult trees (Fig. 9), but spread much more from the stem and have longer tips than the leaves of the mature foliage. The root system is well branched and may be said to have a taproot often 4-5 inches long at this stage. Older trees lack taproots completely.

|
| Fig. 9. Mature sequoia foliage is unique in appearance among trees, Its spreading, awl-shaped leaves overlap each other in a spiral form that gives the branchlets its typical rope-like form. The leaves, which are sharply prickle-pointed, are persistent and shed mostly with the entire branchlet. |
If the readily available soil moisture is plentiful during the second year of growth, the seedling may attain a stature of 8-12 inches and begin to acquire a somewhat roundish or oval crown of many branches. The foliage is now clearly recognizable as the giant sequoia's, being distinct from that of any other North American tree. The roots, still maintaining the juvenile taproot pattern, may now penetrate the soil to a depth of 10-15 inches.
Mature foliage of sequoias consists of awl-shaped leaves, 1/8 to 1-1/4 inch in length on the new shoots. The leaves are closely attached and persistent on the twigs in their basal portions only; the slightly spreading tips are free from the stem and bear sharp prickle points. Their overlapping, shingle-like arrangement gives the stems a rope-like appearance, with the stem completely concealed. The leaves are so persistent that they never fall singly, but are shed intact with the entire twigs to which they are attached after several years of photosynthetic service to the tree. Even as the stem sizes increase in radial growth, the original leaves persist, becoming more widely spaced; eventually the developing bark seems to engulf them, or they are sloughed off with the deciduous bark scales. Both time and shading vitally affect the shedding of the twigs, especially of those in the crown's interior where light is reduced.
In the better sites, upward growth of the leader shoot of seedlings will usually greatly exceed the rate of lateral growth, so that the developing crown becomes narrowly conical and sharply pointed at its tip. The seedlings in poorer sites grow slowly at best and may not outwardly show any growth increment at all. With greater shading, the struggling plant may lose parts of its foliage and thus shrink in overall size, but may maintain a somewhat globular crown for many years. When heavily shaded, the trunks are twisted and contorted into many forms, evidently lacking sufficient light for normal development. Persistent shade brings death to these shade-intolerant trees (Fig. 10).

|
| Fig. 10. Shade-killed young sequoias, Mariposa Grove, Yosemite National Park. Seeded in about 1912, these trees died in the dense shade of white firs which are more shade-tolerant than the sequoias. In the absence of canopy-opening fires, shading is a major death factor to young sequoias. Photo by R. J. Hartesveldt. |
Sapling stage
For several years, the young striplings in the better sites continue to grow rapidly in height and to maintain their narrow, conical crown form, which is now called a "spire-top" (Fig. 11). This sharp-pointed crown may persist for 100 years or more where the light is good, and the growing tip may be as much as 150 ft or more above the ground. The slightly ascending limbs are often dense with foliage, giving the crown a compact form which readily separates it from its associated firs and pines. Seed-bearing cones may also be common at this time, an added means of identifying the sequoia among its associates.

|
| Fig. 11. "Spire-top" sequoia. Young sequoias in the better sites grow upward rapidly and maintain this narrowly conical crown form up to 100 years and more. The larger spire-tops are usually found in locations where sunlight and soil moisture are optimal. Photo by R. J. Hartesveldt. |
At this stage of development, the root system has spread out laterally and the taproot is gone. The depth to which the main root system grows is seldom more than 2 or 3 ft, and ranges outward to 100 ft or more, depending upon soil moisture conditions.
The trunk's diameter may be as much as 4 ft at breast height (4.5 ft above the ground), and the brownish bark may attain a thickness of 1-2.5 inches. If, as rarely happens, a sequoia survives to this stage in the open, its branches may persist nearly to the ground level. Where shaded, the lower branches die and begin to fall away, so that the crown becomes confined to the upper third or upper half of the tree.
The rapid upward growth favors the tree's survival but also invites possible damage to it. Where the tree survives a severe struggle for light by over-topping its competitors, wind and snow may damage its tall, slender stem, either or both breaking off the top or bending it beyond recovery. As it protrudes above the general crown level, it also becomes subject to lightning strikes.
The bark
As the sequoia ages, the grayish scales of the bark begin to slough away and expose the predominantly cinnamon-red bark which gives this tree its characteristically striking appearance on a sunny day. Although its synonym, redwood, may have been derived from the bark color, the more likely origin is the reddish color of the heartwood.
The eventual thickness of the bark is apparently a genetically controlled trait; some specimens seem to have less tightly bound bark scales than others, which results in a sloughing off and, eventually, thinner bark. The species is well-known, however, for the bark's massive thickness, perhaps greater than that of any tree on earth, and certainly a major attribute leading to the tree's longevity. The bark, being soft and fibrous, is a poor conductor of heat, and it contains but little pitch: all these features are important in its renowned resistance to fire damage. As the trunk grows outward, the bark must split longitudinally to accommodate this growth, and over several hundred years the bark ridges' average thickness is about 10 inches. Occasionally, individual sections of bark will exceed 24 inches, and we measured one very limited portion on a tree in Redwood Canyon, Kings Canyon National Park, that was 31 inches thick. The popular literature mentions thickness of 3 ft and more, figures which have never been verified. These represent careless estimates, or perhaps the total thickness of bark on both sides of the tree trunk. Age seems to increase the sloughing off of bark scales, the specimens with the thickest-bark being generally intermediate in age with 8-12 ft trunk diameters.
Another probable genetic trait is the pattern of the bark formed by the flutings and ridges. Perhaps the most basic pattern is a combination of the long parallel ridges and flutings and the reticulate pattern. In this pattern (Fig. 14), bark ridges coalesce or grow together at irregular and often distant points. Less common is the long parallel pattern in which the bark ridges are roughly parallel to each other and the trunk and seldom join each other (Fig. 12). The basic reticulate pattern (Fig. 14) is relatively uncommon, but outstanding in its appearance, and often bears very deeply fluted recesses.

|
| Fig. 12. Sequoia bark pattern showing parallel ridges. although a common pattern, most common is this pattern blended slightly with the reticulate pattern. The Mariposa Tree, Mariposa Grove, Yosemite National Park. Photo by R. J. Hartesveldt. |

|
| Fig. 13. Spiral bark pattern. Perhaps the least common of the major bark patterns, the spiraled form seems to be more common in the Mountain Home Grove than elsewhere. This genetic population suggested that this grove has been separated from other groves for a long period of time. Most spiraled specimens show a right-hand twist; a few of them twist to the left. Photo by R. J. Hartesveldt. |
In some localities, bark with spiralled flutings is common (Fig. 13). In the Mountain Home State Forest, a significant proportion of the total sequoia population has bark of the spiralled variety, most of which spirals to the right. Still other trees drop most of their outer bark scales, subduing the ridges to the extent that the older trees present a smooth-bark surface (Fig. 15). The bark of these trees is relatively thin, and often large segments of the trunk are riddled with sapsuckers' holes.
.

|
| Fig. 14. Sequoia displaying strongly reticulated bark pattern, Redwood Mountain Grove, Kings Canyon National Park. Photo by R. J. Hartesveldt |

|
| Fig. 15. Sequoia displaying thin bark, Grant Grove, Kings Canyon National Park. Some sequoias lose their bark scales readily and present a smooth, thin bark. The protective value of the fire-resistant bark is thus much reduced, a factor which may well explain the scarcity of such trees. Photo by R. J. Hartesveldt. |
Much has been said about the fire-resistance of the sequoia's bark. Very definitely, it is a major factor contributing to this species' longevity. In normal circumstances, there is little or no pitch associated with the bark, which has a fire-resistant property often described as asbestos-like, especially where it is thick. "Asbestos-like" is perhaps a far-fetched analogy, because bark is organic and chars readily with continuous fire from an external source. The loose outer scales will burn with ease, as eloquently described by Muir (1878). The thicker parts of the bark do not hold a flame well, and are seared through only when accumulations of fuel beneath it burn for a long time, or when fire is repeated several times over, as it is especially on the up-slope sides of the trees where fuel accumulates as it moves slowly down-slope by gravity. The very long fire scars that run up trunks are probably largely due to heat from the burning of less fire-resistant adjacent trees such as firs and pines. Fire-scarring often begins in the tree's youthful stages and is repeated over and over again in the event that the tree survives. Cross sections of logged sequoias sometimes disclose fire scars that have completely healed since the damage was incurred. Even remnants of the bark are often embedded in the wood.
Maturity
The value of assigning the term "maturity" to a life stage of this species is questionable. Some trees first produce cones and seeds at the early age of 15-20 years, when they are still slender spires. Where individuals have managed to escape the damaging effects of fires, the rate of vertical growth finally slows in comparision to that of the lateral limbs. Thus the crown, previously pointed at the tip, gradually assumes a broadly rounded form, which many writers have assigned as the mark of maturity. The ubiquity of fire has reduced the perfect round-top, classical in its grace and beauty, to a relative rarity. The Alabama Tree and Galen Clark Tree in Yosemite's Mariposa Grove (Fig. 16), and the Roosevelt Tree in the Giant Forest are excellent examples of this crown form. Where fires intrude through the bark and sapwood, the resulting disruption in the flow of water and minerals to the crown causes a part of the crown, usually the uppermost portions, to die back, which further results in an irregular crown and most often a dead, snag-top (Rundel 1973). Snag-tops are far more common in this stage of the sequoia's life cycle than is the rounded crown form.
Fig. 16. The Alabama Tree, Mariposa Grove, a classical example of the round-top crown form of older sequoias. This crown form indicates a relative lack of fire damage to the base of the trunk. Fire has been a ubiquitous factor in the Sierra Nevada over many years so that this crown form is relatively rare. Photo by R. J. Hartesveldt.The trunks of well-formed trees at this stage may be 10-20 ft or more in diameter above the butt swell. Perhaps the most salient feature of these grand specimens is the trunk's very slight upward taper. This attribute continues into old age, at least in the larger specimens such as the General Grant Tree (see Frontispiece). This characteristic is more prominent in sequoias than in any other Sierran tree. The height of mature specimens varies with both site quality and the degree of fire damage to the trunk's base. Bottomland trees seem generally to be the tallest ones, and specimens of more than 300 ft are certainly not rarities. Despite the rather commonly cited heights of 350-400 ft, there is little to substantiate such claims. One specimen recorded as being 320 ft has been remeasured at 220 ft, and the earlier figure is now regarded as a typographical error. The tallest known specimens recorded by instrument are 310 ft tall, one in the South Grove Calaveras Big Tree State Park (State of California 1924) and the other in the Redwood Mountain Grove, Kings Canyon National Park.
A better figure for the average height of mature specimens is 250-275 ft. Most of these trees bear dead tops, which proclaim a reduction from a former taller stature during a rather indefinite period of years. There is small chance, therefore, that they will exceed their present heights. Occasionally, lateral limbs take over the role of leader shoot and extend upward to greater heights than the original central stem.
The distance up to the first limbs is often impressive too. With the falling away of the lower limbs, the trunks are often bare for 100-150 ft and occasionally more. On the General Sherman Tree, for instance, the largest limb, with a basal diameter of nearly 7 ft, starts at the 130-ft level, and continues in a broad curve for an estimated 125 ft, thus constituting a sizable "tree" in itself.
Apparently, the areal dimension of a sequoia's rooting zone, based upon limited examination, is very nearly an inverse measure of the availability of soil moisture. Along the drainage bottoms where subsurface moisture flow may continue through most of the growing season, lengthy roots are not necessary and may not extend more than 40 or 50 ft away from the trunk. Where the soil is well drained, however, roots grow outward, commonly to a distance of 100 ft and more. Although Muir (1901) reports roots 200 ft long, there is only one other record of sequoia roots growing to that length.
In one study, trenches were dug outward from the base of an almost unburned 12-ft specimen in a well-drained site in the Giant Forest. Roots were followed until they disappeared at a distance of 125 ft from the tree. Assuming a roughly circular pattern of root growth around the tree, the rooting area was calculated at roughly 49,000 ft2, or 1.13 acres. Rooting depths were also measured to calculate the volume of soil occupied by the tree's roots. At the base of the tree, main lateral roots went down nearly 4 ft, but almost immediately proceeded upward until all the feeder roots at the extremities were within a foot of the soil surface. Length and depth measurements established that the volume of soil occupied by roots was about 91,500 ft3.
Within the volume of soil mentioned above, as much as 137,000 gallons of soil moisture could be stored at tensions between field capacity and the wilting point. Although not all of the water within this range is available for plant use, the amount of storage remains impressive. This water is shared, of course, with other plants having their roots in the same volume of soil. Occasionally, during the summertime, soil moisture is replenished to a small degree by thunder showers. But winter snow melt and spring rainfall are the basic sources of supply of soil moisture for giant sequoia growth (Hartesveldt 1965).
The shallow depth of the roots of so large a tree surprises nearly all park visitors. How do such trees remain standing without a deeper anchoring system? Of course, they don't always. The shallow root system is often the tree's undoing and fallen trees expose roots that seldom appear to have gone deeper than about 3 ft. The largest lateral roots are usually a foot or less in diameter although they occasionally exceed 2 ft for a short distance. The trees, it seems, would need to be well balanced to maintain their equilibrium with such a shallow disc of radiating roots to hold them in the soil. Many of them seem perfectly perpendicular, the epitome of a long-enduring record of life. But the number of leaners is surprisingly large, and it remains a mystery what keeps some, such as the huge Grizzly Giant with its 170 lean, from toppling to their death.
Very significant is the number of small feeder roots that branch and rebranch in every direction: they present an enormous surface area for the intake of enough moisture to maintain a crown whose foliage may weigh more than a ton. No one has yet made a surface area calculation, perhaps because of the obvious technical difficulties and labor involved. In the upper 2 ft, the soil contains a striking number of these feeder roots, and they are also abundant at the mineral soil surface, where they catch even the slightest precipitation from short summer thundershowers. Below 2 ft, their numbers dwindle rapidly, although individual roots occasionally penetrate to a depth of 5 ft or more in the drier, better-drained soils. Undoubtedly, the complex intertwining of roots also helps support these huge trees.
Where the zone of soil moisture saturation is near the surface, the roots of bottomland and meadow-edge specimens are considerably shallower. Here, poor oxygenation of the roots prevents or sharply reduces the rate of water passage into the root xylem tissues, and often the roots die.
We will say more about maturity in the chapter on "Life History of the Giant Sequoia."
Maximum size at maturity
Man's insatiable curiosity to determine which is the very largest giant sequoia culminated in a flood of correspondence in the 1920s and 1930s which still bulges the old files of both the National Park Service and the Forest Service. The crescendo of demand for such information led finally to a formal study of contenders for the title. Diameter and height had long been the measurements most commonly cited; but in trees so massive, total volume of the trunk portrays the true size best although other parameters are also useful. Volume, as an expression of size, is a bit more difficult to visualize; it is certainly the most difficult figure to obtain, requiring sophisticated instrumentation and calculation techniques. Accurate measurements were generally lacking during this period of increasing interest in the greatest size.
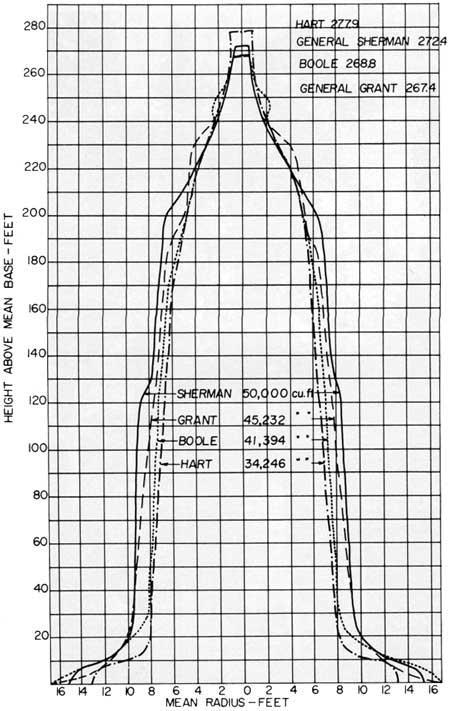
Such was the controversy over which park or which county had the world's largest tree that a group of surveyors, in the summer of 1931, undertook to measure accurately several of the top contenders by means of transits and other surveying instruments. The world's four largest trees were identified in this study and reported by Jourdan (1932) as follows:
| General Sherman | General Grant | Boole | Hart | |
| Height to top of trunk in ft | 272.4 | 267.4 | 268.8 | 277.9 |
| Volume excluding limbs in ft3 | 49,660.0 | 43,038.0 | 39,974.0 | 32,607.0 |
| Volume of burns in ft3 | 350.0 | 2,194.0 | 1,420.0 | 1,639.0 |
| Restored volume in ft3 | 50,010.0 | 45,232.0 | 41,394.0 | 34,246.0 |
| Mean diameter of re stored base in ft | 30.7 | 33.3 | 33.2 | 26.5 |
| Mean diameter in ft at 60 ft | 17.5 | 16.3 | 15.3 | 14.5 |
| Mean diameter in ft at 120 ft | 17.0 | 15.0 | 13.9 | 12.9 |
Of the four above, the General Sherman Tree (Fig. 17) is clearly the largest in volume, mainly because it tapers only slightly. Figure 18, taken from the Jourdan report, compares graphically the taper of the four trees, showing why the General Sherman Tree is the largest even though it has a diameter at breast height (dbh) nearly 3 ft less than the Grant and Boole trees. Another tree not measured in this survey, the Grizzly Giant in Yosemite's Mariposa Grove, is generally considered the world's fifth largest tree, according to transit measurements by park engineers.

|
| Fig. 17. The General Sherman Tree, Giant Forest, Sequoia National Park. Although this is the largest of all trees on earth on a volume basis, it is probably far from the oldest. Situated in a drainageway with a good soil moisture supply, the Sherman's annual growth rings are wider than those of much smaller specimens on drier sites. Estimates place this tree at 2200 years. Photo by National Park Service. |
|
| Fig. 18. Graphic comparison of the dimensions of the world's four largest trees. Despite the fact that the General Sheman Tree is smaller in diameter than the Grant and Boole trees, the slight taper to its trunk provides the added volume that gives it the top honor. Line drawing by Ivan Linderman, permission of Engineering News-Record. |
No one has remeasured the above trees since 1931, so there is a 40-year lag in the statistics. According to our increment borings, taken in 1965, the General Sherman Tree had an annual growth rate of almost a millimeter (1/25th inch), or a radial growth of about 1.5 inches during the 40-year growth period since the study. If added evenly to the entire tree trunk, the net accumulation of wood for that period would be about 1500 ft3, or the amount of wood used in the construction of an average-sized house. This puts the yearly average of new wood production at about 40 ft3, or approximately the volume contained in a tree 1 ft in diameter and 50 ft tall. Such a yearly increment of almost 500 board feet of wood helps substantiate the surprising claim that the world's largest tree may also be the world's fastest-growing tree.
Fused trees and grafted roots
Because of the high density of seedlings in some areas, two or more adult trees growing in close proximity often compete for the same trunk space. When the bark of such trees begin to touch, the pressures created must, in time, destroy the soft, fibrous bark so that phloem, cambium and eventually sapwood (xylem) tissues become fused and continuous around the trees' entire perimeters. The fusion is evident in several specimens where subsequent fires have carved deep caverns and exposed the fused tissues. Here it is not easy to determine just where the wood of one tree ends and that of the other begins. Fusion is basal only, of course, but the fused trunks sometimes continue upward for 20 or 30 ft. In large specimens, the combined circumference may considerably exceed the circumference of the larger single-stemmed trees; some early records of excessively large trees were probably measurements of such dual specimens.
Still another form of tissue fusion evident in the sequoia, but not studied much, is root-grafting. There are several examples of sequoia trunks without crowns that are still "alive." This happens most commonly when a smaller specimen in close proximity to a large sequoia has succumbed to shading, but still continues to "grow" by adding radial wood tissue. One large specimen exists in the Redwood Mountain Grove next to a dense cluster of large, vigorous sequoias. Increment borings indicate that, despite its loss of foliage, the "snag" is still growing at roughly one-third the rate of its nourishing benefactors some 12 ft away.
In other circumstances, where stumps or separated "flying buttresses" of roots are so attached, but with the bark removed to the ground level, the bark has regrown until it covers the entire remnant, giving it an appearance not unlike that of a cypress knee (Fig. 19). Walter Fry in 1903 noted one such specimen in the Giant Forest Lodge area regrowing its bark very near the ground level. The remnant, about 2.5 ft tall, is now completely overgrown with new bark.

|
| Fig. 19. Living remnant of a severed trunk section. Severed from the parent tree by fires long ago, cambial and phloem tissues are fed through root fusion beneath the ground level. Remnant shown in this picture was devoid of bark and living tissues in 1902, is completely covered in 1971. |
Old age
A great majority of sequoia specimens 15 ft and greater in diameter display fire scars that encompass large segments of the trunk's original circumference. Coupled with their great size, this is certainly what gives the older and larger sequoias their uniquely massive appearance and irregular, craggy crowns. Few, indeed, are without a snagged top; the fire scars that caused them to die back are often large and greatly varied in form. These scars undoubtedly give the sequoias the craggy character for which they are best known.
Cavernous fire scars at the bases of sequoia trees have been used for both human living quarters and for stables. Perhaps the one best-known is the Stable Tree in the Mariposa Grove which the U.S. Cavalry used during its administration of Yosemite National Park. The tree fell to its death in 1935. Some specimens have tunnels all the way through the trunk; others are completely hollowed out like a chimney, so that a person standing in the hollowed base can see blue sky through the burned-out upper trunk. A specimen in the Mariposa Grove, the Corridor Tree, has resulted from several fire scars coalescing and forming a corridor between the remnant "flying buttresses" of the swollen base. The intact central section of the heartwood still supports the bulk of the tree's weight.
Perhaps the most remarkably scarred of all sequoias is the Black Chamber in Giant Forest (Fig. 41), which had once a diameter of about 18 ft at ground level. Fires have destroyed 96% of its circumference and most of the trunk's wood tissue. Yet the tree, very much alive, is growing radially at the normal rate of about one millimeter per year. It has a crown of vigorous foliage and is loaded with cones.
No one has ever satisfactorily explained the swollen or buttressed bases of the sequoias. While leaning specimens are known to add compression wood on the down-lean side, perfectly straight specimens often have the largest and most spreading bases (Fig. 20). Furthermore, the suggestion that trees in wet soil and on slopes grow such bases for additional support does not withstand field inspection. Some do and some do not. Many specimens on steep slopes have virtually no swelling in the trunk's basal region, so that the function of the swollen base remains doubtful.

|
| Fig. 20. Sequoia with buttressed trunk, Giant Forest, Sequoia National Park. This condition is common in the species although the one portrayed here is an extreme case. It has been suggested that the buttressed base is a supportive mechanism for trees growing in moist soils or on hillsides. Field evidence for such an explanation suggests that the condition is genetic. Photo by R. J. Hartesveldt. |
The great outward growth of the trunk over the centuries creates another pronounced, though not completely peculiar, feature in this species. Soil, which must be displaced to accomodate the tree's radial growth, is forced outward as the tree expands (Zinke and Crocker 1962), and piles up around the trunk until the tree appears to stand upon a slightly raised pedestal. Such soil becomes somewhat compressed and is referred to as a peripheral pressure ridge. It is most noticeable on the more level sites, where erosional soil movements are minimal. In the absence of fires, the accumulating duff, leaf litter, cones, and bark scales further accentuate these ridges.
Cones and seeds
The seed-bearing cones of sequoias mature at the end of the second year: they are now ovoid, and average between 2 and 3.5 inches in length, and 1.5-2.25 inches in diameter. The cone scales, whose average number is 34, become thickened at the apices and are flattish to slightly recessed, giving the whole cone a solid, compact appearance (Fig. 21). Cone scale variation seems to be genetic: certain trees produce predominantly small cones, while the cones of others are all abnormally large; some have as many as 56 cone scales (Beidleman 1950). One exceptional cone has been found which possesses extra spirals of cone scales for a total of 62 scales.

|
| Fig. 21. Giant sequoia cones, green and closed on the left, dried and open on the right. The cones, which mature at the end of the second summer, do not disseminate seeds while on the tree unless the cone stem is broken or until attacked by small beetle larvae. They are known to have remained attached in a green, growing condition for as long as 21 years. |
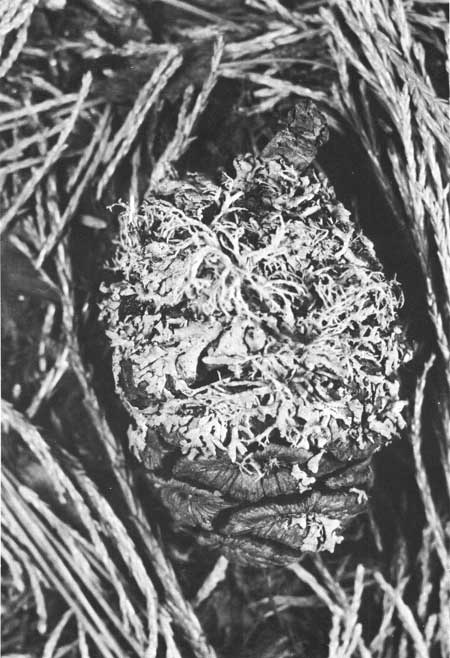
At maturity, the cone scales are forest- to olive-green, and each scale contains a double row of vascular tissue which supports the growth of the expanded fleshy tips. The cones remain green and growing after maturity and continue to grow in size until the lower lips of the scales have become rounded and bulbous. Now they become darker and, after a few years, begin to support a growth of lichens that may in time literally cover the entire cone (Fig. 22).
|
| Fig. 22. Lichen-encrusted sequoia cone. Within 4 or 5 years, growths of lichen begin to appear on some cones. Those cones which remain attached sometimes become so completely encrusted that eventual seed dissemintion is impaired. |
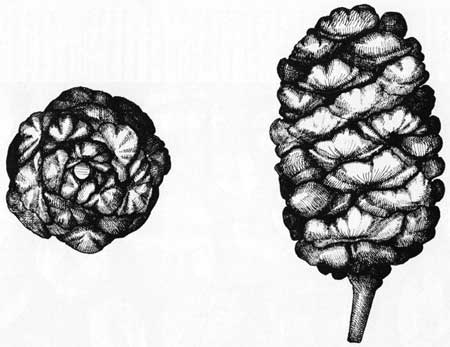
Cone scales are arranged in ascending spiral rows so that, for each species of conifer, there is a definite number of rows around a cone. However, counting to the right and to the left produces different numbers, the ratio thus becoming a diagnostic characteristic. For the giant sequoia, we find three whorls to the left and five to the right when counting from its stem. For the ponderosa pine, the respective numbers are 5 and 8; for the Jeffrey pine, 8 and 13. The numbers are part of the Fibonacci Series, which increases in a predictable pattern from 3 to 5, and from 8 to 13, etc., by adding the first two numbers to obtain the third, etc., beginning with zero.
All plants with an alternate or spiral arrangement of leaves or scales exhibit patterns which fit the Fibonacci Series. Even the flowers and fruits in the head of the sunflower have this arrangement.
Of the many thousand sequoia cones we observed, all but one have displayed the 3:5 ratio (Fig 23). The one exceptional cone has a 5:8 ratio which, while still in the Fibonacci Series, is atypical for this species. We do not understand fully the significance of this precise mathematical arrangement of whorls, so universal in plants. We delight, however, in pondering the unknown biological and physical forces which dictate to the developing cone scales so precise an array.
|
| Fig. 23. Sequoia cone illustrating the 3:5 Fibonacci ratio. Drawing by Ivan Linderman. |
The seeds of the world's largest tree are surprisingly small. At maturity they are mostly 1/8-1/4 inch in length and are perhaps only three-quarters that in width. The tiny embryo is elongated in the direction of the long axis of the seed, which is flat, with straw-colored wings surrounding the embryo and aiding seed dispersal (Fig. 24). Each cone scale produces from four to seven seeds, and cones average about 200 seeds. One pound of seed contains an average of about 91,000 seeds (USDA 1948). More will be said about seeds under Cone and Seed Production, page 88.

|
| Fig. 24. Giant sequoia seeds. produced in prodigious numbers, the seeds of the world's largest tree are surprisingly small—91,000/lb—and proclaim the tenuous nature of this species' reproductive success. Photo by Shirley Fisher. |
Nature of the wood
The sequoia, unlike most of its shade-intolerant associated trees, has soft and brittle wood. Generally, the taller trees in the early seral stage grow upward rapidly, thus maintaining their competitive position in the forest canopy, and the wood must be strong to hold the heavy crown aloft with minimal breakage. Obviously, the sequoia makes up in bulk what it lacks in tensile strength. Its dry weight—18 lb/ft3—is considerably less than that of most forest trees. By comparison, ponderosa pine weighs 29.5 lb; Douglas fir, 32 lb; and canyon live oak, 53 lb/ft3 (Peattie 1953).
The great brittleness of its wood (see Sequoia Lumbering Operations) may have been the sequoias' salvation. The reddish heartwood is made up of short fibers that render it soft and brittle by comparison to the wood of the coast redwood. Hence, it eventually fell into disfavor with the lumbering interests. Its use in lumber products, however, was generally where durability against decay was of greater importance than tensile strength (Noyes 1928). In contact with damp soil, it has few peers and an almost legendary durability. Peattie (1953) claims no signs of decay after 10,000 years.
The brittleness of the wood lay behind its almost unbelievable wastage during the lumbering days when as much as 80% of a felled tree became useless because of breakage. The older heartwood, especially when dry, breaks irregularly across the grain at almost any angle to produce an interesting variety of breakage patterns.
The sapwood is a light yellowish color and is coarse-grained in its early years of growth, when the annual rings are wide. Rings up to 10 and 12 mm wide (approximately 0.50 inch) are common in young trees growing in the better sites. As the tree grows in diameter, the wood cells of the innermost section of the trunk are changed in color by the deposition of chemicals which turn these once-functional cells a pinkish color. This area, now known as heartwood, is largely supportive in its role, and the chemicals which cause its darker color are believed to be the factor that makes the wood so durable.
In the coast redwood, Anderson et al. (1968) have identified chemicals believed to be the specific constituents that make the wood "unpalatable" to both insects and fungi. These have been named sequirin A, sequirin B, and polymeric phenolics. The giant sequoia, whose wood has not been so analyzed, apparently contains similar substances.
The heartwood of the sequoia, with its high tannin content, is not as immune to the activities of decay organisms as earlier writers have led us to believe. This belief is curious, if we consider how commonly heart rot occurs in the numerous remnant stumps in logged areas, and in the form of hollowed trunks of standing trees. Few, if any, studies have been directed at heart rot organisms in the giant sequoia. In fact, Bega (1964) lists only one fungus (Lenzites saepiaria) as a decay agent in this species; yet, for Sequoia sempervirens, he lists several, two of which are common. This suggests that further studies may eventually identify others, such as Fomes sp., in the sequoia. How the hyphae of the decay fungi enter the wood tissue of sequoias is not known.
Decay organisms do not seem to attack all trees: some are sound, while others are affected to a rather large degree. Some specimens are largely hollowed out by repeated fires, which have found the decay-softened wood an easy mark and have thus produced "telescope" trees with openings through the upper part of the trunk. The loss of so much supporting wood tissue weakens the tree and makes it much more susceptible to toppling, undoubtedly the major cause of death of old trees in this species. There is considerable evidence that, when the upper crown dies and the wood dries out, the limbs often become affected by a cubical brown rot. Such evidence has been found at virtually all elevations within the range of the species.
The thickness of the sapwood in relation to the heartwood is directly proportional to the amount of living crown. On trees whose crowns are full, there may be 3 or 4 inches of functional sapwood, but on trees whose crowns are reduced because of large fire scars, lightning, and other kinds of damage to the trunks, the sapwood may be reduced to a narrow band an inch or less in thickness.
Without the tannin content of the heartwood, sapwood on fallen logs usually decays within a few years where conditions are moist. The heartwood remains essentially unchanged for centuries, even where trunks have fallen and remain on moist ground, or are partially buried in it. Although John Muir believed that the wood might last on the ground for 10,000 years, no one knows just what its maximum durability might be under natural conditions. In 1963, three samples of wood were collected from old stumps or log remnants in Sequoia and Kings Canyon National Parks for radio-carbon dating by W. F. Libby and G. J. Fergusson at the University of California, Los Angeles.
Radio-carbon dating is based on the facts that carbon-14 is produced in the upper atmosphere, is radioactive, and has a half-life of 5570 years. When it is incorporated along with carbon-12 into the wood of a tree's trunk, it begins to lose its radioactivity, with half being lost in 5570 years. Thus the ratio of carbon-14 left with respect to the amount of nonradioactive carbon-12 gives a measure of when the wood was formed. If it is the last wood added before a tree dies, the time that has passed since the tree's death can be estimated by this method.
One of three sequoia specimens taken for this purpose from the Circle Meadow area of the Giant Forest was thus calculated to have been dead for approximately 2100 years ± 100 (Hartesveldt 1964). On the basis of such a small sample, we may assume that older specimens probably exist. Muir had hoped that such remnants might be found in forested areas between the extant groves, proving a greater continuous range in former years. None has yet been discovered, however, undoubtedly because the repeated fires would surely have consumed all that may have existed in the past.
Much of the earlier literature credits sequoia wood with low combustibility. While this may be true for the trunks of standing trees that are near full moisture content, the wood generally burns readily when dry. It is virtually without resin except where injury occurs to the trunk (Andrews 1958), and even then the amount of pitch is too slight to be much of a factor in combustion. Peattie (1953) lists the tannin content as the fire-resistant property. Certainly the bark is much more fire-resistant than the dried wood, and its tannin content is known to be high.
The wood of the limbs is especially brittle, probably because the annual rings are narrow. When upper limbs die by any cause such as fire scars interrupting the flow of nutrient solution from soil to crown, the wood dries out and becomes highly susceptible to breakage. Winters are often very destructive to these limbs, which may weigh a thousand pounds or more. Snow accumulation on the crown and winter storms take their toll, and occasionally limbs fall during the summer months, a hazard to visitors walking or sitting beneath the trees.
The wood fibers of the giant sequoia are so short that their value for paper pulp is slight. We do not know of attempts to utilize it for pulp in the United States, but in Hungary, where the tree grows rapidly in the better sites, its pulp has been used experimentally. While the fibers have proven too short to make sufficiently strong paper, further experiments are planned to use sequoia wood fiber as a filler, mixing it with fibers of other trees normally used in paper manufacture.
Longevity
Trees achieve long life in the absence of conditions or factors tending to weaken or kill them. In general, fire, fungi, and insects are the normal decimating agents which kill a tree directly or weaken it, thus hastening its toppling. Most trees have an identifiable age of increased susceptibility to the attentions of insects and fungi which shorten the tree's life span. We do not fully understand these relationships in the giant sequoia, nor can we identify for it an age of susceptibility comparable to that of other trees. The largest and the oldest-appearing trees continue to grow with apparent vigor, putting down annual layers of wood whose volume is about the same as in previous centuries. Indeed, if the giant sequoia remained free of fire damage and could otherwise remain upright, it seems that specimens might well live to the great ages sometimes predicted for them. And in these circumstances of old age, the reproductive capacity of these trees appears undiminished.
A common fallacy about trees is that size is directly proportional to age. The assumption is basically sound, but fails to take into account the very different average annual growth rates dictated by environmental conditions, especially the availability of soil moisture. It is difficult to comprehend the logic sometimes used to estimate great age, or the strength of the contentions proffered to the public. This belief even misled John Muir, who said, ". . . the largest trees are always the oldest and therefore are found upon ridge tops isolated from fire by rocky barrens or by streams" (Wolfe 1938).
Probably no aspect of sequoia life history has been so disputed as its believed antiquity. Diameter has commonly been the basis for age assumptions and they were often calculated from the ring-width of nearby cut trees that were possibly growing under different ecological circumstances. The accuracy of such ratio-and-proportion calculations has been in doubt almost since the first estimates were made. In reference to a calculation by Torrey, a comment was made that "Its enormous size is owing to its continued rapid growth rather than to any very extraordinary age" (Anon. 1854). The suggestion was ignored and subsequent calculations ignored this simple but well-founded explanation.
The exact age of trees is uncertain even from the ring counts of stumps; calculating the age of standing, living trees presents a much more difficult situation. Trees are almost never cut at ground level so that early growth rings are not recorded at stump height. Furthermore, under certain climatic growth conditions, annual rings may not be represented at average stump height or at the height at which increment borings are normally made. Yearly additions to trunk growth typically begin in the crown area and proceed downward until growth is retarded or stopped by autumn climatic conditions. When radial trunk growth is retarded or stopped by a short summer and an early fall, rings at breast-height level may be absent.
Yet, such tenuous age estimates for standing specimens are still common in the literature. Increment borings are possible for short depths only into the trunk of a tree, and the greater the diameter, the less the accuracy of prediction. Claims of increment borings striking the center ring of a large tree should be completely discounted. The standard increment borer is only 15.75 inches long, and specially ordered instruments are only up to 3 ft in length, while ours is only 25 inches long. Furthermore, the chronological and geometric centers of most trees are seldom if ever at the same point. Chances of hitting the chronological center are remote at best, even if an increment borer of that length were available.
For the giant sequoia whose climatic conditions are not greatly variable over a long period of time, missing rings are known to be relatively uncommon. Schmeckebier (1912) states that sequoias never fail to add an annual ring, a statement later refuted by Gillette (1930) who found missing rings in a large specimen. Nevertheless, age-dating by ring count remains more accurate than any other method known for the giant sequoia.
Excessive age figures were derived early in the sequoia's human history, were passed from one writer to the next, and tenaciously held onto, some even surviving until recently. Without a doubt, the simple application of ratio-and-proportion arithmetic yielded many of the earlier figures, ranging from 5000 to 10,000 years. The botanist Willis Linn Jepson pointed out the fallacy of this method many years ago, when he established that a sequoia stump in the North Calaveras Grove, whose age by ring count was 1300 years, would have been 6480 years old by the ratio-and-proportion method of estimation (Lewis 1955). The General Sherman Tree, likewise on the basis of being the largest, has been widely heralded as the oldest living thing on earth.
Muir (1894) reported counting more than 4000 rings on the stump of a large specimen in the Kings River Forest (Converse Basin), a figure widely repeated by others in subsequent writings. As no one has ever been able to locate said stump, or any other with a ring count even close to that number, the report remains unverified. Even the Ellsworth Huntington Expedition into the Converse Basin, after counting rings on more than 450 stumps, failed to locate such a long-lived specimen.
At this writing, 3200 years of age is the oldest count on record. The stump count was made by A. E. Douglass, the well-known dendrochronologist, before 1920. But in spite of such knowledge, some still hold to much older figures for the greatest age, and a recent author claims one specimen to be 6000 years old.
The age of the larger specimens continues to evoke considerable interest. The park visitor often inquires about it, and the answers are not always easy. Age varies greatly with growth conditions within a site. Trees growing where soil moisture is readily available throughout the growing season will produce wood tissue at a rate approaching twice that of a tree the same size in a site where the soil moisture regime is poor.
A relatively recent method of calculating a living sequoia's age is a modification of a method reported by Spurr (1952); it takes into account the tendency of annual rings to become thinner with increasing age while maintaining a rather steady state of wood production. The larger the radius of the trunk becomes, the thinner the rings required to maintain the average growth.
By this method, age is most easily calculated by establishing the basal area of the trunk's cross section at the height at which one or more increment borings are made; then plotting it as a regression curve against time by adjusting the radius as derived from the cores for each 25- or 50-year period back to the innermost ring on the core. During the earlier years of growth, the basal area pattern is curvilinear, but then becomes approximately a straight-line function as the tree becomes older and the photosynthetic increment is evened out over many years (Fig. 25). The basal area pattern in these average circumstances, then, is straight enough in most specimens to allow useful calculations of age that are much more reliable than the older ratio-and-proportion method. With the straight-line assumption, we can establish the growth pattern of a living giant sequoia for the last foot or two of radial growth by one or more short increment borings, which provide the necessary figures to establish a basal area-to-age-slope when graphed. With the slope established, we can use a straightedge to project the basal area line toward the zero basal area level, keeping in mind the early years' curvilinear growth pattern. This better insures that calculations will include the all-important site quality inherent in the steepness of the charted slope. The steeper the slope of the growth pattern, the more rapid the growth, and the more likely for the tree to have optimal environmental conditions. In studies to date, the steepest slopes we have calculated are those of the General Sherman and General Grant trees, the world's two largest living organisms.

|
| Fig. 25. Basal area regression curve showing basic growth pattern of giant saequoia, and used for estimating the age of living specimens. The solid part of the graphed line represents the basal area increment over the period of years represented by the core removed from the tree. The dashed line is a continuation of the straight-line function already plotted. During the earliest years of growth, basal area increases to present a curved pattern and is thus the most difficult part of the curve to assess. Age estimations, however, probably are more reliable than by other methods applied to living, standing trees. Line drawing by Win Stiles. |
To facilitate predictions by this method, 100 complete cross-sectional growth curves were plotted from records taken in the Converse Basin by Ellsworth Huntington in 1911. The derived curves not only portray a vast range of growth conditions but also show that not all the growth patterns have followed the straight-line assumption earlier suggested. A few seem to reflect a continuously improving availability of soil moisture, perhaps as the roots extended outward and intersected more reliable sources of soil moisture. This, of course, complicates the estimation of age. For bottomland specimens, where soil moisture has been more reliable, the growth patterns are fairly steady, and calculations are apparently accurate within 5-10% of the known age.
The results of age dating by this method indicate the need for some rather drastic revisions of sequoia age estimates. According to an average of three cores taken above the butt swell of the General Sherman Tree, this largest of all trees is less than 2500 years old. In 1931, it was cored by Douglass, who calculated its age at about 3500 years by another method (pers. comm.). Estimates of the General Grant Tree, the second largest in the world, now give it less than 2000 years. The Grizzly Giant, fifth largest, is perhaps only 2500 years old, whereas earlier estimates based upon size and its unusually rugged and craggy appearance placed it at 3800 years. Cores which we removed from the Wawona Tunnel Tree after it fell in 1969 indicate that its age was close to 2200 years.
The giant sequoias' reduced age has brought shocking disappointment to many, and a final recognition that the bristlecone pine is truly the king of sylvan antiquity. Actually, this effects a change only in superlatives; the world's largest trees are the world's fastest-growing trees. They are growing on sites of optimum quality.
Refinement of this method of age calculation aims toward establishing a computer program which we hope will permit more accurate age predictions, perhaps within 5% of the age measured by short increment borings.
Meanwhile, 3200 years is the oldest known sequoia record based upon ring count. Findings of false and missing rings have been rare in sequoias, so that the 3200-year figure must be reasonably accurate. That older specimens will eventually be found is a good prospect. Furthermore, assuming that a continuing management program will prevent fire damage and falling, we believe man may eventually discover whether this species actually has an age of susceptibillty to diseases and insects. Such a determination is, of course, many generations in the future.
Sequoia burls
Burls (Fig. 26) of considerable size are far less common on giant sequoia trees than on the trunks of the coast redwood. They occur at varying heights on the trunks up to about 100 ft and may be 3 ft or more in diameter. Unlike the redwood burls, those of the giant sequoia lack the ability to sprout foliage when cut off and placed in water.

|
| Fig. 26. Sequoia burl, Mariposa Grove. Burls are not uncommon on trunks of the giant sequoia, although less so than on the coast redwood. The growths, often as large as a washtub, are now believed to be caused by a soil bacterium. Although sometimes referred to as cancerous growths, they are not known to kill the trees. Photo by National Park Service. |
The cause of burls in sequoia was somewhat of a mystery until Martin (1957-58) conducted experiments in Germany using burls that occurred on the stems of relatively young nursery stock. In these experiments, he isolated a soil bacterium, Agrobacterium tumefaciens (Smith and Townsend) Conn., which when injected under clinical conditions into the stems of uninfected specimens, always produced burls. We do not yet fully understand the mode of introduction into native specimens at many feet above ground level. However, an early account relates that a falling sequoia splashed mud and stones up to 100 ft on the trunks of neighboring trees. Conceivably, soil bacteria could have entered the stem through wounds caused by the flying rocks. This same bacterium is recorded as a crown gall organism on both giant sequoia and coast redwood seedlings (Bega 1964).
These limited observations on the occurrence of burls or galls do not preclude several other causes such as genetic, physiological, or other factors.
Other large trees
Man has favored the giant sequoia because of its size and age, symbols of strength and durability, but many claims have been made for other great trees. Apparently, these other species have suffered similar inadequate measurement and, like the sequoia, have shrunk by today's standard measures.
Actually, comparing diameters of tree trunks raises some difficult questions: what is accepted as a trunk, and is the trunk a single stem or the fusion of several? Past comparisons have been notably careless, or perhaps intentionally vague. We know or strongly suspect some of the largest trees to be fused, yet we must give credit to their remarkable size even though comparisons with single-stemmed specimens are not really valid.
Other species of large trees, some either taller or greater in diameter than the giant sequoia, are listed below. The sequoia, as a single-stemmed tree, remains undisputedly the largest tree on earth in trunk volume, and only the bristlecone pine has been proven older.
Coast Redwood. Sequoia sempervirens (Lamb.) Endl.
Family: Taxodiaceae.
Native habitat: central to northern California, into southern Oregon.
The only really close contender for volume size with the giant sequoia is its cousin, the coast redwood. Ecologically, the latter is quite different from the sequoia and grows considerably taller in its foggy habitat along the Pacific Coast. Many specimens considerably exceed 300 ft, and one specimen in the northern range is nearly 370 ft tall. With trunk diameters commonly 10-12 ft but rarely exceeding 16 ft, total volume is considerably less than in the giant sequoia. The oldest recorded specimen is just over 2200 years of age, hardly a contender for the longevity record, but yet no youngster.
Baobab. Adansonia gregorii F. Muell.
Family: Bombacaceae.
Native habitat: Australia.
Although the largest specimens have 30-ft diameters, they are seldom more than 40 ft tall; a few, however, have been recorded up to 60 ft. The claim that this strange tree is more than 5000 years old seems debatable, as it produces no annual rings. The wood is reportedly very soft.
Douglas fir. Pseudotsuga menziesii (Mirb.) France.
Family: Pinaceae.
Native habitat: western North America; California north into British Columbia, east to the Rocky Mountains.
Although the Douglas fir has been occasionally recorded as the world's tallest tree, it probably ranks third or fourth, following the coast redwood, eucalyptus, and giant sequoia. The Douglas fir grows up to 220 ft, far less than record measurements of the other species mentioned. Diameters rarely exceed 10 ft, and claims of 17 ft are unsubstantiated. The tree is important as timber; no species in North America is cut in greater quantities than the Douglas fir.
Ceiba or Kapok. Cieba pentandra (L.). Gaertn.
Family: Bombacaceae.
Native habitat: Central America.
This handsome tree with huge basal buttresses grows to more than 12 ft in diameter above the butt swell, and up to 150 ft tall. Measurements around its buttressed bases have undoubtedly caused the tree to be claimed as the world's largest.
Eucalyptus, Mountain Ash, or Peppermint Gum. Eucalyptus regnans F. Muell.
Family: Myrtaceae.
Native habitat: Australia and Tasmania.
This species of eucalyptus overtops the tallest giant sequoia if its earlier height measurements are accurate. On Mount Baw Baw in Gippsland, Australia, is a specimen 326 ft tall, and one near the Styx Rivet in Tasmania is 322 ft tall. The larger diameters are from 6 to 9 ft. Claims of 450-ft heights are fairly common, and Robinson (1882) places the tallest at 600 ft.
Dragon Tree. Dracaena draco L.
Family: Liliaceae.
Native habitat: Canary Islands.
Much of the earlier literature describes one specimen as impressively large. The tree which blew down at Teneriffe in 1868 was 45 ft in circumference, or 14.3 ft in diameter, and 70 ft tall. Claims that it was 6000 years old have no support in scientific literature.
Tule Cypress or Ahuehuete. Taxodium mucronatum Ten.
Family: Taxodiaceae.
Native habitat: Mexico.
A trunk of this species at El Tule, near Gaxaca, Mexico, has a circumference of 162 ft, or a diameter of 51.6 ft. This diameter is considerably greater than that of the largest giant sequoia, but botanists generally believe that this specimen is composed of several fused trunks, and that the comparison is therefore invalid. Another famous specimen, the Montezuma Tree, in Mexico City's Chapultepec Park, has a diameter of 14.3 ft and is 200 ft tall.
Kauri Pines.
Family: Araucariaceae.
Native habitat: New Zealand and Australia.
Queensland Kauii. Agathis robusta (C. Moore) F. M. Bail. Native to Queensland, Australia. Maximum diameters are recorded as 7-9 ft, and heights up to 160 ft.
Agathis palmerstonii F. Muell. Diameters up to 10 ft.
Agathis australis Salisb. One specimen, now destroyed, at Mercury Bay, New Zealand, measured 23.8 ft in diameter, and the first branch was 80 ft above the ground. Estimates, on unknown basis, put its age at 4000 years.
Fitzroya. Fitzroya cupressoides (Molina) Johnston.
Family: Cupressaceae.
Native habitat: Swampy areas in southern Chile.
Specimens are reported up to 15 ft in diameter and up to 240 ft tall, with some age estimates exceeding 1,000 years.
Jaquitiba Tree. Carinaria excelsa Casar.
Family: Lecythidaceae.
Native habitat: Brazil.
The Jaquitiba, a relative of the Brazil nut tree, is recorded as having diameters up to 23 ft and being 150 ft tall.
Bo-tree, Peepul, or Pipal Tree. Ficus religiosa L.
Family: Moraceae.
Native habitat: India and Ceylon.
As its scientific name suggests, this tree has religious significance. Vishnu, an Indian deity, was supposedly born in the shade of a bo-tree. Like its relative, the banyan, it usually has numerous trunks. One specimen on the Narbudda River in India measures 2000 ft in circumference around its 650 trunks. Another specimen was brought to Ceylon from India in 188 B.C., so that it was 2161 years old in 1973.
Sweet or Spanish Chestnut. Castanea sativa Mill.
Family: Fagaceae.
Native habitat: Europe, from Great Britain to Spain and southern Italy.
The largest specimen recorded was at the foot of Mount Etna in Sicily. Named "Castagno del Cento Cavalli" (Chestnut of a Hundred Horses), it was still alive in 1935, but is now dead and destroyed. The Encyclopedia Britannica mentions its having a circumference of 190 ft in 1780, and Chambers Encyclopedia of 1836 gives the circumference as 204 ft. Tiemann (1935) and William Steere of the New York Botanical Gardens, in a personal communication, report that this otherwise amazing trunk was again due to several sprouts fusing into one. Other large specimens are reported in Sicily, and England claims a tree with a 15-ft diameter in Gloucestershire.
We have not included here other species of impressive proportions, such as the sitka spruce and Western red cedar, because no claim was ever made for them as the world's largest. (See Menninger 1967 for an excellent account of the world's larger trees.) Certainly, the issue is far from closed, although the giant sequoia seems to have no serious competitor for the title of largest in volume.
| <<< Previous | <<< Contents >>> | Next >>> |
chap2.htm
Last Updated: 06-Mar-2007