|
OLYMPIC
The Evolution and Diversification of Native Land Use Systems on the Olympic Peninsula A Research Design |

|
Chapter 3
THE REGIONAL ENVIRONMENT AND FOOD RESOURCES OF THE OLYMPIC PENINSULA
by Randall Schalk and David Yesner
Physiography and Climate
The Olympic Peninsula includes a land area of roughly 6,000 mi2 bordered by the Pacific Ocean on the west, the Strait of Juan de Fuca on the north, and the Hood Canal on the east. The interior of the Peninsula is dominated by the Olympic Mountain massif with individual peaks rising to elevations of more than 2400 m (ca. 8,000 ft). Most of the Peninsula landscape shows dramatic evidence of glaciation. Sharp peaks and rugged ridges separated by steepwalled valleys are characteristic in the mountains. Pleistocene valley glaciers that extended nearly to the Pacific have left U-shaped valleys through which rivers such as the Hoh, Queets, and Quinault flow. The Strait of Juan de Fuca and Hood Canal are fords that were cut by Pleistocene ice.
The mountainous core of the Peninsula has a radial drainage pattern centered about the south to-north flowing Elwha drainage. There are about 50 glaciers in the Olympics which amount to an area of 65 km2 (25-30 mi2) that is permanently covered by snow and ice (Danner 1955).
Along the west side of the Peninsula there is a coastal lowland that extends inland for a distance of 16-24 km (10-15 mi). Along the north central margin of the Peninsula, the coastal lowlands are compressed to a width of less than 8 km (5 mi) but from Port Angeles east to the northeastern corner of the Peninsula this coastal plain widens somewhat to 16 km (10 mi) or more. In contrast to the western Peninsula, the mountains along the east side drop precipitously to the Hood Canal and are drained by rivers that have minimal sections of low gradient in their lower reaches. Moving southward along the Hood Canal, there is virtually no coastal plain until one reaches the Skokomish at the southern end of the Canal. The south face of the Olympic mountains is drained by a number of tributaries to the Chehalis River such as the Wynooche and Satsop whose lower reaches flow through an extensive lowland.
Those rivers that originate in glaciers high in the Olympics tend to have stability in discharge throughout the year. In contrast, smaller, independent streams that only receive tribute from low elevation areas tend to be intermittent in flow. These flow characteristics result from the highly seasonal pattern of precipitation.
Most precipitation falls between October and May and summers tend to be relatively dry. Below elevations of roughly 750 m (2500 ft), virtually all precipitation falls as rain. Precipitation increases steeply with increased elevation. Prevailing winds move in from a southwesterly direction and lose most of their moisture on the windward slopes of the Peninsula. A rainshadow created on the leeward side of the Olympic mountains is centered about Sequim, Washington where annual precipitation is only 15 in (375 mm). This is the dryest coastal area in the Northwest and yet it is juxtaposed within a few miles of areas that receive in excess of 200 in (5000 mm) of precipitation annually (see Figure 3.1). This region is clearly one of the most climatically heterogeneous areas of North America. In general, the Olympic Peninsula is exceptionally complex physiographically and this complexity is manifested in marked climatic, biotic, and ethnographic variability.
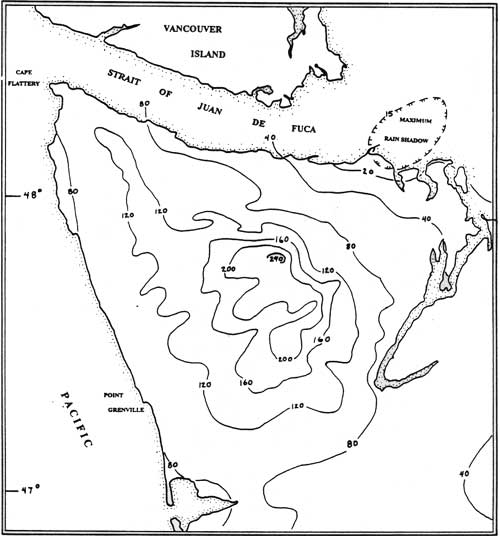
|
| Figure 3.1 Mean Annual Precipitation (inches) on the Olympic Peninsula: 1930-1957 (redrawn from Phillips and Donaldson, 1972). |
The Olympic National Park encompasses a substantial range of the Peninsula's environmental variation. Both the windward and leeward portions of the central mountain massif are largely within the Park. Although lowland reaches of most of the Peninsula's rivers lie outside of the Park, a narrow arm of the Park extends nearly to the mouth of the Queets River embracing a lowland riverine environment that has considerable archaeological potential. On a smaller scale, a stretch of the lower Quinault also falls within the Park boundaries. Finally, the coastal strip section of the Park encompasses an 80 km long stretch of coastline facing onto the Pacific Ocean. This part of the Park is a high energy coastline and the oceanographic features of this area are discussed and contrasted with the other sections of the Peninsula coast later in this chapter.
The bedrock of most of the interior of the Peninsula's interior is composed of basalt, sandstone, shale, slate, and phyllite (Tabor 1975). The basalt bedrock occurs in a roughly horseshoe-shaped distribution around the northern, eastern, and southern portions of the Peninsula (Tabor 1975).
Vegetation
Prior to the advent of the timber industry in Washington, most of the Olympic Peninsula was covered by dense primeval forests. Some of the world's highest primary biomass occurs in the lower river valleys of the western portion of the Peninsula where summer fogs reduce evapotranspiration to produce rainforest conditions (Heusser 1974:1548). The largest living examples of certain conifers are found on the western Peninsula as a result of combined effects of heavy precipitation and long growing seasons. The rivers, lakes, small prairies, and areas above timberline were the only interruptions in this otherwise continuous forest.
The Pacific coastal forest of the Peninsula's lowlands is composed of western hemlock (Tsuga heterophylla), western red cedar (Thuja plicata), and Sitka spruce (Picea sitchensis). When these climax conifer stands are displaced by fire, blow-downs, or disease, seral communities appear that are composed mainly of red alder (Alnus rubra), black cottonwood (Populus trichocarpa), bigleaf maple (Acer macrophyllum), and willows (Salix sp.). These broadleaf stages give way to Douglas fir (Pseudotsuga menziesii), grand fir (Abies grandis), and western white pine (Pinus monticola) as succession progresses towards climax conditions. Fluvial action also creates early seral stages so that the various stages of forest succession may be seen as one moves up from modern river floodplains over terraces of increasing age (Fonda 1974). An association of alder, black cottonwood, bigleaf and vine maples (Acer macrophyllum, A. circinatum) typically occurs along flood plains of streams (Heusser 1977:285).
The elevational zonation of forest types can be divided into lowland, montane, and subalpine associations (Fonda and Bliss 1969). The lowland forest extends up to elevations of between 550-600 m (1800-1980 ft; Fonda and Bliss 1969). Up to this elevation, virtually all precipitation falls as rain while both snow and rain occur at higher elevations.
The montane forest is dominated by Pacific silver fir (Abies amabilis) and western hemlock in the western Olympics. On the drier east slopes of the Olympics, however, the montane forest contains Douglas fir and western hemlock as dominants and this forest type may extend up to 1400 m (4,620 ft) in some areas. The influence of forest fire is evident in the east slope forests and accounts for the importance of Douglas fir there.
At elevations of about 1,075-1,125 m (3550-3700 ft) in most areas, the transition from montane forest to subalpine forest occurs. This zone of transition in forest types, like that between the lowland and the montane forests, relates to a seasonal snowline—in this case associated with early summer season (Fonda and Bliss 1969). The melting of snow upslope in the spring generally reaches an elevation of about 1,100 m or the lower edge of the subalpine forest by June.
The subalpine forest type extends up anywhere from 1,400 to 1,650 m depending upon aspect. Pacific silver fir and mountain hemlock (Tsuga mertensiana) are the dominants in the cool, moist mountains of the western Olympics. In the drier eastern Olympics, subalpine fir (Abies lasiocarpa) forest types occur.
Understory vegetation throughout the forest types varies in ways that are undoubtedly of direct relevance to the distribution of forage for herbivores such as elk and deer. For example, the montane forest has an exceptionally well developed shrub layer but a poor herb layer (Fonda and Bliss 1969). Within the subalpine forests, west slope Pacific silver fir/mountain hemlock type has a well developed tree-sapling-shrub layer but a poor herb layer. East slope Abies lasiocarpa forests have an understory that is poor in saplings, shrubs, and herbs. In general, it appears that increasing snow accumulations at higher elevations contribute to an overall suppression of understory moving along an elevational gradient through the forests.
At its upper margin the subalpine forest grades into parkland and meadows. This transition occurs between 1,100 and 1,450 m elevation. Based upon differences in vegetation composition, eight different meadow types have been distinguished (Kuramoto and Bliss 1970).
Fire Ecology and Forest Vegetation
Forest fires modify natural vegetation in very important and strongly patterned ways. In considering the characteristics of fire in this ecosystem, we must consider two basic types—that caused by lightening and that ignited by humans either deliberately or inadvertently. A sizable body of information is available on the former whereas very little is known about the latter for this region.
Lightening caused fires tend to be of greatest consequence with the occurrence of three conditions: 1) several months of drought, 2) ignition source in the form of a thunderstorm, and 3) winds blowing from the east which are typically dry and warm in the summer (Agee and Flewelling 1983). The co-occurrence of these conditions is relatively unusual to judge from the historic records but there is tree-ring evidence to suggest that these conditions were more common at certain intervals in the past. During intermittent periods of drought in past centuries, lightening fires were much more common (Agee and Flewelling 1983).
Given these critical variables, one would expect that there would be contrasts in the importance of fire between the west and east sides of the Olympics and this, in fact, is the case (Fonda and Bliss 1969; Pickford et al 1977; Agee and Flewelling 1983). Lightning fires occur in highest frequencies on the dry east side of the Bailey Range and are relatively rare west of there (Pickford et al. 1977). On the east side, seral forests predominate while on the west side climax forests are widespread. Major fires apparently burned most of the east side forests about 300 years ago (Fonda and Bliss 1969:294). In the lowlands, Douglas fir is the major seral species while in the subalpine zone, subalpine fir develop after fire events (Fonda and Bliss 1969:294).
Besides the east-west contrast in fire frequency, there is also an important elevational pattern in the occurrence of lightening fires. Most of the fires historically in the Olympic National Park have occurred above 3,000 ft elevation. As much as 77% of the lightening fires in the eastern part of the Park were at elevations above 3,000 ft (900 m; Pickford et al. 1977:31). It would seem that the concentration of lightening fires at higher elevations is partially responsible for the maintenance of the subalpine parks and meadows. These areas are heavily used as summer range by elk and deer and the quality and quantity of such range may have been improved during drier intervals in the past. Major fire episodes in the Cascade Range since 1300 A.D., for example, have been correlated with reconstructed drought intervals (Hemstrom and Franklin 1982).
A second source of fire is human ignition. There is an extensive literature on the controlled application of fire by hunter-gatherers around the world (c.f. Lewis 1982) and the use of fire to maintain early successional vegetation is well documented in the Northwest as well (Norton 1979). However, ethnographic data pertaining specifically to subsistence burning on the Olympic Peninsula are scarce. It is known that there were a number of prairies in certain river valleys on the western Peninsula. Singh (1956) lists 18 named prairies, most of which are in the lower portions of the watersheds. It is clear that there were additional prairies in this area or portions of the Peninsula that were not reported by Singh. [1] Olson (1936:52), for example, states that "Meadows are rather infrequent but there were a sufficient number along the creeks and on the margin of the lake [Quinault] to insure an adequate supply of camas to vary the meat diet" Inspection of USGS quad sheets revealed three additional prairies on the western Peninsula that are not mentioned by Singh (1956). All of the prairies for which locations are known are listed in Table 3.1 by drainage; locations of these prairies are shown in Figure 3.2.
TABLE 3.1
PRAIRIES OF THE OLYMPIC PENINSULA
| Location/River Valley | Prairie | Map# |
| Moclips Valley | Moclips Prairie Baker Prairie | 1 2 |
| Quinault Valley | Mounds Prairie Chow Chow Prairie O'took Prairie |
3 4 5 |
| Whale Creek | Moses Prairie | 6 |
| Queets Valley | Unnamed Prairie | 7 |
| Quillayute Valley | Little Prairie Quillayute Prairie Forks Prairie Maxfield Prairie Tyee Prairie Beaver Prairie Small Prairie Elk Prairie Hoh Prairie Middle Prairie Last Prairie |
8 9 10 11 12 13 14 15 16 17 18 |
| Ozette Valley | Ahlstrom's Prairie Manny's Prairie Roose's Prairie |
19 20 21 |
| Sequim Vicinity | Sequim Prairie | 22 |
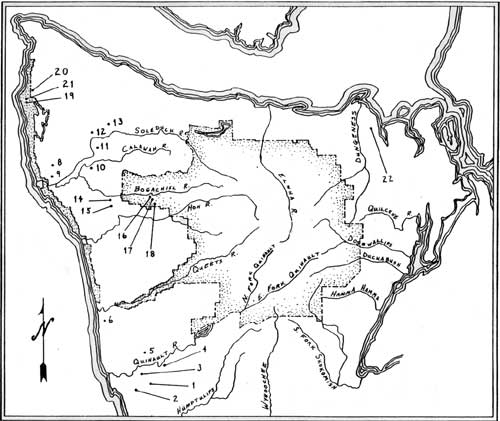
|
| Figure 3.2 Prairies of the Olympic Peninsula (redrawn from Singh 1956). (click on image for a PDF version) |
Why prairies were seemingly concentrated on the western side of the Peninsula is not at all clear but the only known exceptions to this pattern seem to be near Sequim. Petersen et al. (1983) describe Sequim Prairie as being 800 ha in area and, owing to the extreme summer drought characteristic of this area, it may owe its existence to different factors than the prairies of the western Peninsula. Gunther (1927:272) mentions a prairie on the "upper Elwha" but details about the specific location or size of this prairie are not mentioned.
Prairies appear to have been important plant collecting locations that were maintained by the Indians through the use of fire. The Indian agent on the Quileute Reservation, Albert Reagan, suggested that burning was done to promote forage for deer, especially bracken:
The burning of this fern year by year was what kept up the "prairies" of the peninsula and extended these areas. The Indians burned the ferns for the purpose of clearing out the prairies so they could shoot the deer and elk when they came to feed on the young fern sprouts. They also dug the roots of these ferns, pounded them to a pulp, mixed it into a sort of dough and baked bread from it, which they claim was pretty good bread (1934:56-57).
Singh (1956:43) states that these prairies were important to the Indians of the western Peninsula and that they were deliberately burned on a regular basis:
Each family had an "occupancy right" in a great prairie such as O'Took Prairie or Quileute Prairie. The family burned over its part of the the prairie in the spring so that dead ferns would be destroyed, giving way to camas. Men hunted elk, deer, smaller animals on the prairies.
Reagan's conclusion regarding the reason for burning the prairies has been challenged on the grounds that bracken is not an important forage species for deer (Norton 1979).
Norton (1979) points out that a common association of plant species is found on prairies in settings that range from rainshadow settings to very wet areas of western Washington. She has identified 46 species of plants that were used by native peoples of western Washington and included on this list are virtually all of the major sources of carbohydrate that were available in this region.
Berries (Rubus sp.), camas (Camassia quamash), and bracken fern (Pteridium aquilinum) are the plant foods ethnographers most commonly identified as the objective of controlled burning in western Washington. All of these species are not only fire resistant but are actually enhanced by burning (Suttles 1951; Norton 1979). Although not confirmed in recent archaeological investigations, Reagan (1917) reported the remains of ancient earth "oven mounds" containing burned camas roots, fruits, and clams at various locations along the western Olympic Peninsula.
Plant Food Resources
A rather diverse range of plant resources was exploited by the people of this region (Reagan 1934; Gunther 1988). Given the overall structure of this ecosystem, however, edible plant resources are not sufficiently abundant to have served as dietary staples. The significance of plants may go beyond quantitative importance though and it is clear that the roots, berries and sprouts used by native people in the Northwest provided valuable sources of minerals such as calcium, magnesium, iron, zinc as well as ascorbic acid (Norton et al 1984). The use of camas occurred primarily in the southern part of the Northwest Coast but the distribution of this resource was limited to openings in the forest and places that are moist in the spring but dry by summer (Turner and Kuhlein 1983). Because plant resources like camas provided virtually the only sources of carbohydrate in a predominantly meat diet, such resources may have been important for their role in improving metabolic efficiency (Speth and Spielmann 1983).
Different species of huckleberries occur from near sea level up to the subalpine zone. The fire maintained subalpine parklands of the eastern Olympics would seem to offer the optimal habitat for huckleberries. Nonetheless, the abundance of huckleberries in the eastern Olympics seems significantly below that of areas in Washington's southern Cascades. In addition to huckleberries, there are a number of other edible plants that occur in the high country of the Olympics. These would include cow parsnip (Heracleum lanatum), miner's lettuce or spring beauty (Claytonia sibirica), and various tubers. Given the labor costs of collecting most of these resources in any quantity, however, it is doubtful that most of these were more than very minor supplements to the staple food resources.
Land Mammals
In most temperate zone coniferous forests, ungulates tend to dominate the terrestrial faunal biomass effectively exploited by hunter gatherers. Historically, the Roosevelt elk (Cervus elaphus roosevelti) and black-tailed deer (Odocoileus hemionus columbianus) were the only ungulate species of the Olympic Peninsula. There is presently no evidence that any other ungulate species have been present in this region since late glacial times. It is possible that mountain goats (Oreamnos americanus) and other species were present in this region prior to 10,000 yrs B.P. but became extinct during the Holocene (Lyman 1988). This point will be discussed at greater length in a later chapter but the important fact is that the terrestrial fauna of the Olympic Peninsula might be considered an "island fauna". In other words, the montane fauna of the Olympics are separated by substantial habitat barriers to other mountainous areas. The general ecological principle that islands tend to have less faunal diversity than continents applies well to this region.
The Roosevelt Elk
The Roosevelt elk is the largest and most abundant ungulate of the Olympic Peninsula and the Olympic herds represent the largest population of Roosevelt elk along the entire west coast of North America. Due to drastic reductions of elk primarily from overhunting in the late 1800s, Olympus National Monument was created in 1905 for protection of this species. Despite protection for elk in the area that was to eventually become Olympic National Park, areas outside the Park have been dramatically modified due to settlement and logging. Historic settlement was concentrated in the lowlands where extensive areas of prime habitat for elk were eliminated. Logging, on the other hand, probably increased capacity of the region to support elk (Taber 1980; Starkey et al. 1982). It is estimated that there were between 25,000 and 40,000 animals on the Olympic Peninsula in the 1800s (Skinner 1936), only 2,000 in 1905 (Starkey et al. 1982), 6,000-7,000 in 1945, and 15,000 in 1982 (ibid). Assuming that these figures are reasonably accurate, it would appear that any beneficial effects of logging have not compensated for other adverse factors.
The Roosevelt elk differs from the Rocky Mountain Elk (Cervus elaphus nelsoni) in several respects. The former is larger; average weight ranges for Roosevelt bulls are 700-1,000 lbs and for cows 400-700 lbs (Schwartz and Mitchell 1945). There are differences in coat coloration and antler development (ibid.). Roosevelt elk have a lower reproductive rate (Starkey et al. 1982). Habitat preferences are significantly different with Rocky Mountain elk primarily grazers that prefer open forest conditions whereas Roosevelt elk prosper in dense old growth forests and make extensive use of browse plants. The old growth forests are not structurally uniform but instead are quite patchy with numerous openings, significant areas of early seral vegetation, and a variety of plant community types. Areas maintained in early seral stages by fluvial action such as deltas, floodplains, and beaver meadows are believed to have supported the highest densities of elk in the coastal forests (Raedeke and Taber 1982; Jenkins and Starkey 1984:645).
Roosevelt elk browse primarily in winter and in summer both graze and browse in about equal proportions (Schwartz and Mitchell 1945). Western hemlock, western red cedar, Douglas fir, vine maple (Acer circinatum), alder, huckleberry (Vaccinium sp.), and salmonberry (Rubus spectabilis) are the principle browse species (Schwartz and Mitchell 1945; Skinner 1936). Most of the plants listed here apparently occur in greatest abundance at lower elevations and progressively decrease with higher elevations—especially above 3,000 ft (900 m; Skinner 1936:256). There is a scarcity of elk forage in the mid-elevation, montane forests. Grasses are concentrated in the the wide valleys of the western Peninsula and in the subalpine meadows but are nowhere abundant in the region. For about one or two decades after forest destruction (e.g. forest fire or, more recently, logging), there is a lush growth of grass.
Given the substantial contrast between the two elk subspecies, it is less surprising that Roosevelt elk vary inversely in abundance with black-tailed deer throughout the Olympic Peninsula. Taber (1980) has suggested that Roosevelt elk of the Olympics vary positively with precipitation and that black-tailed deer vary inversely. The wetter and denser forests of the western slopes are dominated by elk whereas the dryer, more open forests of the east side are dominated by deer. Schwartz and Mitchell (1945:305) estimated that 85% of the elk on the Peninsula were concentrated in the western river valleys. The mild, moist climate of this area is conducive to higher primary production and forage availability than elsewhere on the Peninsula (Taber 1980). During the summer, the headwaters of the Quinault, Queets, Hoh, and Elwha are gathering areas for large numbers of elk (Schwartz and Mitchell 1945). Skinner (1936:256) estimated that "probably one-fifth as many elk live on the north as on the west side" of the Olympics. He also suggested that there were only a few bands in winter on the east slope but that these mingle during the summer with west slope bands (ibid.). Apparently due to the dry, open character of vegetation in the northeastern Peninsula, there are no elk there in summer (Schwartz and Mitchell 1945). South slope river valleys such as the Humptulips and the Skokomish are probably intermediate to the east and west slopes in terms of game abundance.
Elk on the Peninsula include both migratory and non-migratory herds. The migratory herds winter at elevations between 450-600 m (1500 and 2000 ft)—well below the winter snowline at 750 m (2500 ft). The winter ranges of migratory and non-migratory elk in the Olympics are illustrated in Figure 3.3. Migratory elk move into the mountains during the summer where they may gather in large herds to feed in subalpine meadows and parkland above 4500 ft. This upslope movement follows the retreat of snow and results in the greatest yearly dispersion of elk during the summer. Towards late summer, however, there is apparently a tendency to form larger bands.
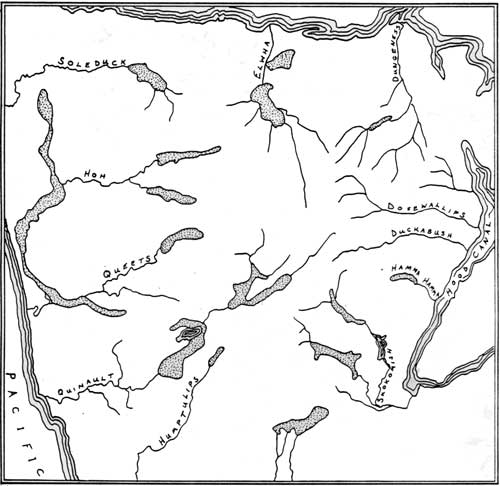
|
| Figure 3.3 Winter ranges of Roosevelt elk on the Olympic Peninsula (redrawn from Schwartz and Mitchell 1945). |
Non-migratory elk, while they make minor adjustments in habitats selected at different seasons, remain in the lower drainages throughout the year. During the winter season, they are found at elevations somewhat below the migratory elk (below 450 m/1500 ft). Presumably, non-migratory elk were once far more extensive throughout the Olympic Peninsula. The areas of the Peninsula having the widest coastal plain, especially the western lowlands, must have once had large numbers of elk. Where the lowland zone is of minimal width, as it is along the Hood Canal side of the Peninsula, only small elk herds could have been supported and these probably would be migratory due to the foraging constraints of the very limited lowlands.
Offsetting the reduction in non-migratory elk habitat due to development, Taber (1980) suggests that logging in mountainous areas has benefited the migratory populations. In this regard, some authorities on the Olympic elk view migration patterns in Roosevelt elk as at least partially if not completely a post-contact development (c.f. Starkey et al. 1982); human settlement in the lowlands is postulated to have driven elk into higher elevation forests (Bailey 1936, cited in Starkey et al. 1982). In the western Cascades, Roosevelt elk were concentrated in lowland river valleys and prairies from northern California to southern British Columbia (Murie 1951; Schoen 1977:33). Historical records suggest an absence of Roosevelt elk in the Cascades north of Mount Rainier (Schoen 1977:33). The archaeological record of the high elevation interior sections of the Olympics is obviously of particular importance for its potential to lend greater temporal perspective on this issue. It is clear that elk made use of the high meadows of the Olympics at the time of the 1890 O'Neil Expedition (Wood 1976:176).
The riparian hardwood vegetation zone provides a major source of winter and spring season forage for elk in this environment and can be a limiting factor in regulating reproductive success (Jenkins and Starkey 1984:645). The wider, U-shaped valleys of the western peninsula are of particular importance as winter habitat due to the quantity of these gallery hardwood forests.
Groups sizes of elk vary throughout the year from dispersal in small bands in spring and early summer to aggregation in late summer before the rut and again in late winter. During the fall rutting season, group sizes are again small. Herds of more than 50 individuals are frequently seen in the subalpine and alpine meadows during the late summer (Taber 1980). If results of studies of transplanted elk in the Cedar River watershed of the western Cascades (Schoen 1977) can be generalized to the Olympics, herd size in closed forest environments is considerably smaller than in the more open forest of the Rocky Mountains.
Black-tailed Deer
Black-tailed deer occur throughout the Olympic Peninsula but, as mentioned above, are most abundant in the dryer areas. Taber (1980) suggests that deer numbers vary negatively with precipitation on the Peninsula and, therefore, are the dominant ungulate in the leeward Olympics. The mature, closed-canopy coastal forests are characterized by low quality and low quantity forage for deer (Taber 1961). Deer densities in this habitat type may be as low as 1-5/mi2 (Taber 1961:229). Where openings in the forest occur, however, densities may rise to 40-50/mi2 (Taber 1961:230). Except for the prairie areas of the windward Peninsula, openings in the forest occur mostly on the eastern slopes of the mountains due to the much greater importance of fire there. Fire has markedly beneficial effects for deer forage, deer density and reproductive rates.
In regions such as the the Olympics where significant snow accumulations occur annually at higher elevations, black-tailed deer migrate between summer and winter ranges along an elevational gradient (Taber 1961). There are, as with elk, both migratory and non-migratory populations of deer in the Olympics. The distribution of migratory and resident deer winter ranges is analogous to that described above for elk. As was also suggested with elk, logging has provided much more extensive forage for deer and this factor may have contributed to greater numbers of migratory deer.
Other Land Mammals
There are a number of other mammals native to the region but which are not likely to have played important roles in subsistence in prehistoric times. These would include wolf, beaver, marmot, otter, mink, fisher, rabbit, squirrel, skunk, and raccoon. While some of these smaller mammals may have been taken occasionally either for food or for fur, there is no evidence or ecological reason to suggest that use of these resources was more than incidental to other primary subsistence activities. Exploitation of these resources would be expected to be embedded in procurement of more important resources and, therefore, of limited consequence in conditioning the location of most archaeological remains. This conclusion must be qualified by the recognition that the economics of the fur trade era may have resulted in changes in the ways in which terrestrial fur bearers were exploited.
Black bear may be somewhat of an exception to these statements and this species appears to have played a greater role than the smaller mammals mentioned above in certain areas; the upper Queets Valley, for example, was known ethnographically for having an abundance of bears and a number of specialized bear hunters (Olson 1936). Judging from the accounts of the Press Expedition through the Olympics (Wood 1967), the number of bears in certain parks and meadows in the mountains was substantial in the summer.
Grizzly bear were not native to the Peninsula in historic times but Elmendorf's Twana informants maintained that an occasional grizzly might be seen in berry collecting grounds south of the Skokomish (Elmendorf 1960). These were characterized as "strays" from the Cascades that might be seen "once or twice in a generation" (ibid).
Ethnographic Data on the Hunting of Land Mammals
The ethnographic literature tends to be lacking in details about procurement of terrestrial resources such as elk, deer, and bear but there are some data worthy of brief summary. The Makah are reported to have hunted land animals rarely and apparently had very limited access to game in their home range (Swan 1869). There is some reason to believe that elk did not often frequent areas near the shoreline (Swan 1869; Olson 1936) but the reasons for this are not entirely clear. The Quileute utilized bow and arrow, spears, and deadfalls for game (Pettitt 1950:10).
Quinault subsistence apparently involved a greater dependence upon large game than did either Makah or Quileute (Olson 1936:4). Seasonal movement to the mountains by entire families often occurred in the late summer (ibid:41). Male efforts focused on hunting from a "semi-permanent camp of poles and brush" while females and children concentrated on drying the meat or gathering berries, basket grasses, and bark (ibid:41). Bow hunting was apparently the primary technique in the mountains but dogs were used for hunting in the river valley (Olson 1936:42). Singh (1956:72) makes a similar distinction between bow hunting in the mountains and the use of dogs for driving game in lowland areas. It seems reasonable to infer from Olson's account that seasonal hunting trips into the mountains in the summer were made by hunters from upriver villages. Transport costs would seem prohibitive for downriver groups.
Some groups of Twana seem to have been more dependent upon hunting than any other Peninsula groups. The Vance Creek band of the Twana, located on a tributary to the south fork of the Skokomish River, are described as completely "inland" in their economic orientation (Elmendorf 1960:56). Fall communal elk drives are reported for Skokomish and other Twana groups on Hood Canal (ibid:58). Elmendorf (1960:30) notes that place names were poorly developed for areas further away from the watercourses and this might be interpreted as evidence that travel into the deep mountains was not commonplace. Though the bow is listed as the main hunting device used by the Twana, a wide variety of other less commonly used techniques are mentioned (e.g. surround, spears, nets, pitfalls, snares, and deadfalls; Elmendorf 1960:86).
Klallam hunting is described as being limited to a few specialized hunters who visited the mountains (Gunther 1927:215). There are no explicit details to confirm whether hunting in the mountains in such instances means the foothills or the high elevation parks and meadows. Also, there are no clues provided as to what season of the year hunting occurred in the "mountains".
Marine Resources
Although Olympic National Park comprises some 4,000 m2 of the Olympic Peninsula, only a small percentage of that area lies in the section adjacent to the Pacific Coast. Prehistoric seasonal rounds undoubtedly included areas that lie outside the boundaries of the Park on the Peninsula so that any regional study of land use must consider the whole Peninsula and not be constrained by modern property boundaries. Also, it is significant to note here that the Park lies close to the coast in some areas, particularly on the Juan de Fuca Strait. A proper understanding of the relationship of marine resources to prehistoric archaeology in the region, therefore, requires discussion of marine resources on both the east and west coasts of the Peninsula. The western portion of the Peninsula will be emphasized, however, since the only portions of the park that front directly on saltwater are along the outer coast.
The eastern and western coasts of the the Olympic Peninsula are distinct in terms of physiography, oceanography, and climate, and this has led to significant differences in the types of fauna found in both locations. The western Olympic coast is a "rugged", high energy coastline characterized by rocky headlands, sea-stacks, and offshore rocky islands. Because of strong onshore winds and because of a strong south-to-north pattern of longshore drift, sandy areas are often restricted to small coves and pocket beaches. In contrast, along the eastern portion of the Peninsula, gravel and sandy beaches predominate. This area is characterized by a low energy coastline, as wave energy tends to dissipate within the sheltered waters leeward of the Olympic Peninsula. While estuary zones are not extensive anywhere on the Peninsula, the outer Olympic coast has no more than 2 km2 of estuary. On the Hood Canal side, there is a fairly extensive estuary and delta on the Skokomish River with smaller ones on rivers such as the Dosewallips, Duckabush, and Hamma Hamma. The Strait of Juan de Fuca coast is essentially intermediate in these features, since its western portions are subject to intense wave action but its eastern portions (east of the Elwha) are relatively protected. Thus in terms of physiographic and oceanographic features of the Olympic coastline, the area may be subdivided into major zones: the Outer Coast, and the Inner Coast, with the latter including the shore of the Strait of Juan de Fuca, and the western shore of Hood Canal.
Oceanography of the Outer Coast
On the outer coast of the Peninsula, nutrients that enrich the food chain derive from two primary sources—the Columbia River and the Strait of Juan de Fuca. In the first case, the increased nutrient levels result from a plume of sediments derived from the runoff of the Columbia River (Peterson 1972; Anderson 1964), while in the second case, they result from the entrainment of nutrient-rich subsurface waters by the estuarine-like circulation of the Strait of Juan de Fuca (Stefansson and Richards 1963, 1964; Winter and Anderson 1975). The effect of these systems is reflected in higher levels of nitrates, phosphates, and silicates in the coastal zones adjacent to these two areas during mid winter. The northward-drifting plume of the Columbia river brings concentrations of nutrients as far north as the Strait of Juan de Fuca while the discharge of the latter ranges as far south as the central Washington coast. In spring and fall, an additional source of nutrient enrichment results from wind-induced upwelling which occurs along the outer Washington Coast. One large upwelling system occurs offshore from Grays Harbor, while a smaller but intensive system occurs near Umatilla Reef, offshore from Cape Alava. This results in significantly cooler offshore waters along this portion of the coast than in the area south of Grays Harbor. It also results in a phytoplankton bloom, particularly in the spring, as the nutrients diffuse upward into the photic zone (Stefansson and Richards 1963, 1964; Anderson 1964; Peterson 1972; Lewin et al. 1975). Tidal mixing serves to increase the strength of the phytoplankton blooms, particularly along the northwest Olympic Peninsula coast. Similar phenomena occur along the southwestern coast of Vancouver Island, as a result of tidal mixing and the entrained plume of the Strait of Juan de Fuca (Mackas et al. 1980). All of the nutrients introduced by these processes are then available as the base of a rich marine food chain, including invertebrates (e.g. shellfish), fish, sea mammals, and sea-birds.
As indicated above, the outer coast can be characterized as a high-energy environment. During winter in particular, average wave heights exceed 5 m as much as 15% of the time; maximum wave heights may reach 8 m or more (Loehr and Ellinger 1974; Phillips 1972). In contrast, waves in the Strait of Juan de Fuca exceed 5 m no more than 4% of the time, even in mid-winter. Wind velocities follow similar trends as noted above. Tidal ranges are also significantly greater on the outer coast, averaging from 1.8-1.9 m on the outer coast and from 1.2-1.4 m east of Crescent Bay. The western portion of the Strait of Juan de Fuca, at Clallam Bay for example, shows intermediate tides of about 1.5 m. In addition to the strong tidal currents, high wind velocities, and large waves that characterize the region, there is a strong south-to-north trend of longshore drift. Thus, any sediment has little opportunity to accumulate. Although there are numerous pocket beaches, cliffs and rocky shorelines are the rule. These cliffs are derived from Tertiary rocks and sediments uplifted from 30-100 m above the modern coast; most sand deposits present in the area are derived from erosion of these cliffs rather than from accretion in the coastal zone. Although there are tidal platforms, these tend to be stripped of sediment through the process of longshore drift. Furthermore, the continental shelf (as defined by a 100 fathom or 183 m contour) is relatively narrow in this area, averaging ca. 15-20 km in width, and offshore rocky platforms tend to dip steeply, particularly in the northern part of the area.
Within the outer coastal area, there are some important environmental differences that deserve mention. North of the Hoh River, the coastline tends to be more complex, with more headlands and rocky islands. This has consequences for the aggregation of sea-mammals and sea-birds in this region, as will be discussed below. To the south of this region, from the Hoh River to Point Grenville, the coastline tends to be straighter, rocky headlands and offshore islands are fewer, and a larger number of sandy beaches are found. Erosion of coastal bluffs and greater accumulation of sand and silt in the coastal zone result in these differences. Although south of the Outer Coastal Study unit, a third coastal zone may be identified in the area from Point Grenville south to Grays Harbor, in which broad, gradually sloping sandy beaches come to predominate and rocky headlands disappear.
These differences in coastal landforms are associated with differences in the offshore continental shelf. To the south of the Hoh River, the continental shelf ranges from 35-50 km in width, while to the north of this area it gradually decreases from ca. 20 km to no more than 5 km in width off Cape Flattery (Rau 1973; Tabor 1975). Offshore gradients tend to be much steeper in this area because of the intrusion of the Juan de Fuca submarine trench. These differences in geological features among the zones of the outer coast have had significant consequences for the development of the resource base that supported prehistoric human societies in the region.
Oceanographic Features of the Inner Coast
The essential differences between the outer and inner coasts of the Olympic Peninsula result from the differences in oceanographic features of these areas, as well as certain distinctions in the terrestrial landscape. In general, the inner coast is a relatively low energy coastline in comparison with the outer coast. Nevertheless, the outer portion of the Juan de Fuca Strait (west of the Elwha River) retains many of the features characteristic of the outer coast, such as a highly eroded shoreline with rocky headlands. East of the Elwha River, significantly lower energy predominates, and more extensive sandy beaches, and spits (e.g. Ediz Hook, Dungeness Spit) tend to accumulate to a larger extent. Further to the east are found still more protected waters of Hood Canal. The Olympic mountains shelter the Hood Canal from the westerly winds. In reality, therefore, the inner coast may be divided into three sections: the outer portion of the Juan de Fuca Strait, west of the Elwha River, the inner portion of the Strait, including the northwestern region of Puget Sound, as far south as the mouth of the Hood Canal; and the western side of the Hood Canal itself. These three subdivisions also roughly correspond to the areas occupied in historic times by three major ethnic groups: the Klallam on the south shore of the Strait of Juan de Fuca, the Chimakum in the Port Townsend region, and the Twana on the Hood Canal.
Oceanographic features of these three sections of the inner coast show some apparent trends as one moves from west to east. During winter (October-March), prevailing winds may be easterly as well as westerly in the outer Juan de Fuca Strait, and they average in excess of 20 mph. In the eastern portion of the Strait, they average between 10-20 mph, while east and south of Port Townsend average winds rarely exceed 10 mph (Loehr and Ellinger 1974). As indicated above, while wave heights exceed 5 m as much as 15% of the time off the outer coast during winter, such waves occur no more than 4% of the time in the Strait of Juan de Fuca, and less than 1% of the time in Hood Canal. These variations in wind conditions have important consequences for the intertidal shellfish resources which are discussed below. Tidal ranges, also, show some variation, being greater than 1.8 m on the outer coast, 1.5 m in the outer portion of the Strait of Juan de Fuca, and 1.2 m in the inner portion of the Strait and the Hood Canal.
In spite of the differences in physical oceanography on the inner and outer coast, the productivity of both regions is equally high. While the Strait of Juan de Fuca may lack some of the enriching effects of local upwelling during summer along the outer coast, it contains an "estuarine-like" circulation which greatly increases the primary production of the coastal zone, particularly during the fall (Stefansson and Richards 1963). This occurs as deeper waters enter surface layers through a process termed "entrainment". During fall, levels of nitrates, phosphates, and silicates develop in the surface waters of the Juan de Fuca region, and mixing of these waters in the photic zone leads to pronounced reductions in oxygen saturation as well as nitrate and phosphate "anomalies" in this region (Stefansson and Richards 1963). In the Hood Canal and Puget Sound region to the east, upwelling is even less pronounced, but true estuarine conditions occur because of the fiord-like nature of the basin. The result is a longer annual period of high productivity.
The primary productivity of the Sound is due to intensive upward transport of nitrate by the estuarine mechanism. Algal growth in the open waters of the central basin of the Sound is dominated by a number of intense blooms beginning in late April or May and recurring throughout the summer. Rarely, and only briefly, does nitrate become exhausted. The phytoplankton production rate in the central basin of Puget Sound is about 465 g c/sq m/yr (Winter et al. 1975:139).
These levels of primary production exceed those found to the north in the Strait of Georgia, where primary productivity averages ca. 120 g c/sq m/yr (Parsons et al. 1970).
Macroinvertebrate Resources of the Outer Coast
The major relevant features of the littoral ecology of the outer Peninsula coast include high tidal range, strong wave action, and an unusual shoreline which gives little opportunity for the development of sandy substrates appropriate for the development of shellfish beds. Pocket beaches that do exist tend to be small in size and are dominated by sand and gravel, frequently lacking in silt-sized particles. Because of the relative uniformity of the outer coast, the abundance and diversity of invertebrate resources is relatively low in this region, particularly for large, edible molluscan resources often preferred by human groups occupying coastal zones. Shellfish available for human consumption are those that have adapted to high-energy, rocky habitats. As one moves further south, however,—particularly to the south of the Hoh River region—this pattern begins to change, with sand beach forms increasing in abundance and the total diversity of available invertebrate resources thereby increasing as well.
The outer coastal region has relatively greater tides which create a potential opportunity for longer collection periods for shellfish and other intertidal resources by native inhabitants of the region. The steepness of the offshore gradient, however, particularly in the northern portion of the area, results in a reduction of the area in which human groups can collect such resources. This reduction in the width of shellfish habitat probably more than offsets any benefits from increased tidal range and perhaps reduces the overall efficiency of littoral collecting strategies. In addition, the steepness of the offshore gradient and the high wave energy has led to a substantial zonation of intertidal and subtidal organisms, such that different species are less likely to be collectable from the same zone (Rigg and Miller 1949). Because of the nature of semi-diurnal tides in the northwestern Peninsula region, there are pronounced differences between successive high and low tides in any given 24-hour period. Finally, the strong seasonality of the upwelling system off the western Olympic coast would have decreased both the abundance and diversity of invertebrate resources available during the winter period. Taken together, all of these factors lead to the expectation that shellfish would not have been emphasized in aboriginal subsistence on the outer coast of the Peninsula. However, it is important to note that in some areas, for example near Cape Alava, where a combination of the offshore upwelling system and the extensive rocky intertidal platform increased the richness of the littoral zone, better conditions for shellfish collection were available.
Intertidal and subtidal communities of the western Peninsula region have been discussed by Paine and Levin (1981), Dayton (1971), Nyblade (1979), Lubchenko and Menge (1978), and Dalthier (1984). A general conclusion that may be distilled from these authors (c.f. Wessen 1982) is that four major shellfish associations exist in the western Peninsula region, three of which predominate in rocky, high-energy zones. They are as follows:
(1) Sea Mussel-Gooseneck Barnacle (Mytilus californianus-Mitella polymerus) association. This association predominates in the zone below mid-tide (i.e. the lower intertidal to subtidal zone). It colonizes wave-swept rocks in high energy zones. Dayton (1971) notes that seas-mussel "mats" of up to 25 cm thickness may develop as large number of individuals utilize threads to attach themselves to bare rock surfaces. Within these zones, however, competition between individuals is greater, so that the largest are often found in the most seaward portions of the intertidal zone or in the subtidal zone. Also found in this lower tidal zone are other high-energy resistant species such as a wide variety of limpets (Acmaea spp.), barnacles (Mitella spp., Balanus spp.), and giant chitons (Katharina tunicata). The mussels, limpets, and chitons are all relatively easy to harvest, although the former greatly predominate in numbers. Barnacles, however, are more difficult to separate from the rocks, and the nutrient yield is usually lower. In general, however, the biomass and abundance of edible seafood for humans is substantial in this high-energy zone.
(2) Bent-nose Clam-Mossy Chiton (Macoma nasuta-Mopalla muscosa) association. This community is found in moderate wave-energy zones associated with wave-cut rock platforms, as occur near Cape Alava. Species comprising this association frequently are found in intertidal pools; they usually occur as individuals rather than as aggregates. Furthermore, most of the species associated with this community are relatively small, low-yield individuals. Some non-shellfish invertebrates, however, are common in this zone (e.g. the Rock Crab (Cancer productus) and the North Pacific Octopus (Octopus dofeini).
(3) Little-neck clam - Butter Clam - Horse Clam (Protothaca staminea - Saxidomus giganteus - Tresus capax) association. This association develops in sandy beaches which do manage to develop in high energy zones, usually in pocket coves or similar protected areas. Unlike the bent-nosed clam - mossy chiton association, the dominant species here are large, relatively high yield edible species, and where they are found they may be locally abundant. However, areas of their occurrence on the outer coast are generally small and highly localized.
(4) Fingered limpet-Sitka Periwinkle (Acmaea digitalis-Littorina sitkana) association. This association is restricted to the "splash zone" at the nearshore extreme of the intertidal area. These species again tend to be only locally abundant but the overall distribution is significantly wider than that of the bivalves of association (3).
In addition to the above, it is worthwhile to note that a fifth association is occasionally found in the northern part of the area, but increases in importance in the sandier zones to the south of the Hoh River, coming to dominate in the area from Point Grenville south to Grays Harbor. This is the razor clam - Bodega Clam (Siliqua patula - Tellina bodegensis) association. It is associated with low gradient sandy beaches fully exposed to the surf. Razor clams occur in the lower intertidal zone where their ability for rapid digging permits them to occupy sandy substrates that are constantly changing as a result of water action. Although quantities of razor clams sufficient for commercial harvest occur at Kalaloch and Taholah, these are areas of comparatively minor production (Schaeffer 1939). The major razor clam concentrations along the Washington coast are from Copalis southward (ibid).
Marine Invertebrates of the Inner Coast
The result of this high primary productivity is a high secondary productivity, well studied in terms of benthic assemblages in Puget Sound (Lie 1974). Intertidal life forms have also benefited from this high productivity. As a result of the quieter waters and greater sandy deposition characteristic of these environments, and the highly productive estuarine conditions, intertidal invertebrates such as burrowing shellfish fare quite well in this area. Three taxa found only in sandier areas of the northern outer coast are much more prevalent in this region: the littleneck clam, the butter clam, and the horse clam. Another species only rarely found on the northern outer coast, also occurs here in significant numbers: the basket cockle (Clinocardium nuttallii). Mussels were present in this area in relatively limited numbers and these tend to be the bay mussel (Mytilus edulis).
Within the Peninsula region, the Pacific oyster (Ostrea lurida) is now restricted to the inner coastal regions—mainly Hood Canal (though formerly present in Willapa Bay on the southwestern coast of Washington). The oyster's restriction to this area and its high productivity (in certain areas of Hood Canal and Puget Sound) may be attributed to a number of factors related to habitat requirement, particularly temperature and salinity. Although the Pacific oyster was at one time distributed from Baja California to Alaska, the temperature conditions of the Washington coast (in the center of the range) were ideal for oyster growth and reproduction. Within that region, oysters thrive best in areas in which temperature conditions reach 13-18° C in mid-summer (the temperatures required for spawning and reproduction), but which do not fall too close to freezing during the winter. These conditions are met in the sheltered basins of Hood Canal and western Puget Sound, where midwinter temperatures average 6-9° C and mid-summer temperatures average 18-20° C. In addition, these areas have the reduced salinities and sandy environments necessary for optimal oyster habitat (Schaefer 1939; Brittell et al. 1976).
Another species of hardshell clam common to the inner coastal region of Hood Canal and western Puget Sound, but not present on the outer coast, is the geoduck (Panope generosa). This very large clam produces a higher caloric return relative to other shellfish, but considerable energy must be invested to harvest it. Three other species of burrowing clams exist in the area today, but were unknown in precontact times: the soft shell clam (Mya arenaria), the Quilcene oyster (Crassostrea virginica), and the bay scallop (Pecten inidians). In general, burrowing clams thrive in the sandier, quieter environments of this region. Compared to the outer coast then, there is a greater diversity of shellfish available from this region; twenty species of shellfish are commonly found in the region (Nyblade 1979). Within this region, furthermore, strong east-west differences are found in shellfish assemblages; Nyblade (1979), for example, notes that at Pillar Point, on the outer Juan de Fuca Strait, the assemblage is dominated by high-energy resistant forms such as sea mussels, chitons, limpets, and barnacles, while at Beckett Point near Port Townsend, the littleneck clam - horse clam assemblage dominates, along with other sand-dwellers such as basket cockle.
Ethnographic Data on Shellfish Use
Swan (1868) provides some information on shellfish use by the Makah Indians of the Cape flattery region:
Although a variety of bivalves is found, they do not abound as they do in the bays further up the [Strait of Juan de Fuca], and do not form a common article of nutriment, except that mussels of the finest description cover the rocks about Cape flattery and Tatooche (sic) Island, and are eaten whenever the Indian appetite craves them, or whenever the breakers of the Pacific are sufficiently quiet to permit a search. These are either boiled or roasted in the ashes, and are very delicious cooked by either method. Barnacles, crabs, sea slugs, peri-winkles, limpets, etc., furnish occasional repasts. Scallops, which are found in the bays of Fuca Strait, are excluded in their list of food.
Additional details on exploitation of invertebrate resources of rocky outer coasts come from aboriginal groups in other regions to the north that are similar to the area held by the Makah. For the Manhousat of western Vancouver Island and the Skidegate Haida of the Queen Charlotte Islands, Ellis and Swan (1981) were able to document relative preferences, seasonal use, locations of exploitation, and harvesting and preparation methods used. According to their data, the following frequently used species of shellfish were found widely distributed in local rocky habitats: the sea mussel, the goose barnacle, the black katy chiton, the giant red or gumboot chiton, the green sea urchin, and the giant red sea urchin. "Occasionally eaten" species from the same habitats included barnacles (Balanus spp.), limpets, black turban snails (Tegula), and bay mussels. Several other species were used that were restricted to sandier areas. Of these, the larger species (e.g. the butterclam) were considered sufficiently large, tasty, and abundant in specific locales to make special expeditions to sandy beaches in order to obtain them. They also rendered themselves most readily to storage for winter consumption (by drying).
Other species, which were relatively rare in the rocky environments of the outer coast, were not often exploited; these included basket cockles, horse clams, Pacific oysters, and Northern abalones (Haliotis kamschatkanz). These species were more commonly found (and used) on the southern and eastern sides of Vancouver Island (or the northern and eastern parts of the Olympic Peninsula). Often these were known to inhabit subtidal sandy areas, but were not often exploited in those zones. One species, the littleneck clam, was taken only incidentally to the harvesting of butterclams. Two species not present in the area were obtained by trade, primarily for decorative purposes: scallops and dentalia. Finally, small gastropods, such as the olive shells, dire whelks, or dogwinkles, were not eaten, probably because of their small size, but many of these were important for ceremonial purposes. Perhaps the clearest picture that emerges from the data, however, is that specific localities were visited to obtain particular species. This was true of most of the commonly exploited species: the California mussel, the goose barnacle, the black katy chiton, and the giant red chiton. The black katy chiton and butterclams were considered sufficiently important to establish overnight camps for exploiting them at more distant, productive locales.
These accounts of the use of rocky coastlines are distinctly different from the account of shellfish utilization among the Quinault Indians in the southern part of the Outer Coast (Olson 1936). Although the Quinault also utilized sea mussels and chitons to some degree, they placed emphasis on the abundant shellfish in sandy beaches that were characteristic of that region. The razor clam was the most important shellfish to the Quinault and was collected in a number of areas south of Point Grenville (Olson 1936). Of secondary importance were the butter clams and Pacific oysters. Crabs were also utilized to some extent (Olson 1936).
In Gunther's (1927) report on the Klallam, she notes the use of horse clams, basket cockles, and butter clams in particular. Of secondary importance were bay mussels, which were only gathered in winter, and "china shoes" (chitons). Other species (oysters, geoducks) were not prevalent in this area, becoming common only to the east of the Elwha River.
For the Twana, along the western portion of the Hood Canal, the most important species were considered: (1) littleneck clams; (2) butter clams; (3) basket cockles; (4) horse clams; (5) Pacific oysters; (6) geoducks; and (7) bay mussels. It was said of the latter that "although abundant [they] did not seem to have been an important food source"; and they were "regarded as unfit to eat at certain seasons" (Elmendorf 1960:58, 124).
Sea Mammals of the Outer Coast
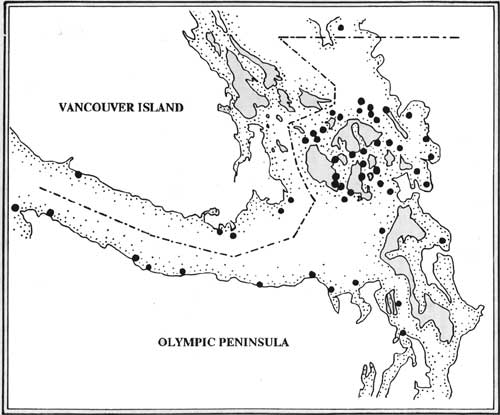
Like the sea-birds, many sea-mammal populations vary seasonally, as a result of feeding habits or long-distance migration patterns. This is true of a number of species important to prehistoric human occupants of the outer Olympic coast: the northern fur seal (Callorhinus ursinus), the Steller sea lion (Eumetopias jubatas), the California sea lion (Zalophus californianus), the northern elephant seal (Mirounga angustirostris), and a variety of baleen whales, including the gray whale (Euchrichius glaucus), the fin whale (Balaenoptera physalus), the sei whale (B. borealis), the Minke whale (B. acutorastrata), and the humpback whale (Megaptera novaeangliae). Many of these species come close to the coast across the continental shelf during their migrations (Figure 3.4).
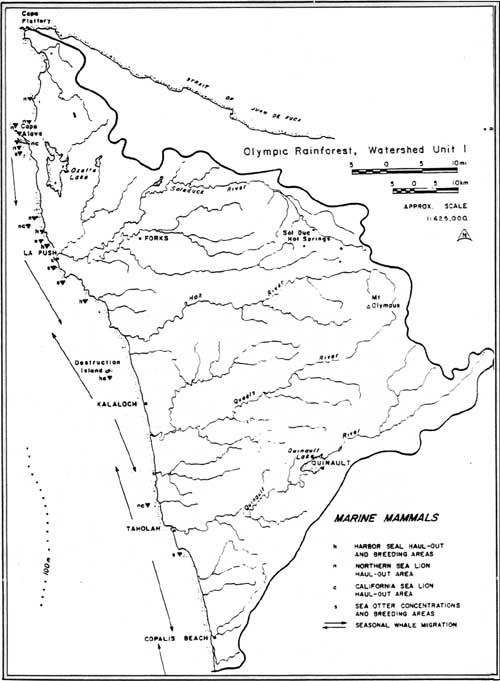
|
| Figure 3.4 Marine mammal concentration and migration areas of the northwestern Olympic Peninsula (from Proctor et al. 1980). (click on image for a PDF version) |
(1) Northern Fur Seals are present in the region in large numbers during the winter, on their way to and returning from their breeding grounds in Alaska. Their first appearance occurs in June while northbound. Fur seals are strictly pelagic off the Washington coast, only rarely venturing into inside waters of the Strait of Juan de Fuca. They do not haul out anywhere within the study area, only occupying rookeries in California and Alaska. During migration along the Washington coast, their path essentially follows the margin of the continental shelf, where they feed on a variety of fish such as anchovy, eulachon, salmon, herring, lamprey, stickleback, and rockfish (Fiscus and Kajimura 1965, 1966). The fur seal migration route comes close to the coast only in the area between Umatilla Reef (off Cape Alava) to Cape Flattery (Everitt et al. 1980). Until at least the 1920s, thousands of fur seals moved along the Washington coast as part of this annual migration.
(2) Northern or Steller Sea Lions occasionally winter off the coast of northwestern Washington and the adjacent coast of southwestern Vancouver Island, before returning to California during the breeding season. They are present in largest numbers between October and April (late fall to early spring), when they feed on a variety of fish such as whiting, walleye, herring, salmon, and various flatfishes (Everitt et al. 1980). Along the northwestern Washington coast, about 500 animals haul out in five main locations: Tatoosh Island, Jagged Island, Split Rock, Spike Rock, and Umatilla Reef (Kenyon and Scheffer 1961). Two additional haulouts occur north of the Hoh River, and a third north of the Quinault River (Figure 3.5). Southwestern Vancouver Island and the mouth of the Strait of Juan de Fuca provide an additional 15 haulout areas, including Race Rocks and Sombrio Point (Smith 1972; Everitt et al. 1980).
|
| Figure 3.5 Locations of known harbor seals haul sites along the Strait of Juan de Fuca (from Everitt et al. 1980). (click on image for a PDF version) |
(3) California Sea Lions are present in the period from late fall to spring (October to May), after the conclusion of the breeding season in California. Tatoosh Island and the Quillayute Needles off the outer Washington coast are important haulouts for these animals; another haulout exists north of the Quinault River (Figure 3.5). Larger numbers appear to haul out in the Strait of Juan de Fuca (e.g. at Sucia Island). Still larger numbers haul out at nine locations along southwestern Vancouver Island, particularly on Race Rocks (Everitt et al. 1980). Up to about 500 animals currently haul out off the Washington coast, while about 500 more are found on Vancouver Island. The major food items of this species are whiting and cephalopods.
(4) Northern Elephant Seals, the largest of the north Pacific seals, presently occur in small numbers off the Washington coast. They are apparently present primarily during the non-breeding season, although sightings have occurred at all times of the year. Little is known about the ecology of this species in the northern part of its range, although principal foods are known to include fish, cephalopods, and benthic, neritic, and demersal prey (Everitt et al. 1980; Cowan and Carl 1945).
(5) Gray Whales are present in large numbers primarily during spring migration (during March and April), as they migrate northward to their summering grounds in the Chuckchi Sea. Nearly the entire population (originally estimated at more than 15,000 individuals) migrated past the Washington coast during this period. A smaller return migration occurs during late November to early January, and because of the imprecision in timing of these migrations some stragglers are found along the coast at all times of the year. Gray whales are usually encountered as individuals or in small pods of 2-3 animals. This is a nearshore-migrating whale, found primarily in shallow, coastal waters, usually passing within a mile of the shore, often in sight of land. They are found along the outer coast to the north of Grays Harbor, migrating past Tatoosh Island as they move northwest, passing along the west coast of Vancouver Island before continuing on their way to Alaska. Some animals also venture into the mouth of the Strait of Juan de Fuca, particularly into Clallam Bay. Gray whales do not feed during migrations.
(6) Minke Whales are present along the Washington coast as a part of their summer range. During winter, they move into lower latitudes off California and Mexico. However, some animals tend to be present throughout the winter. They prefer more protected waters, and are therefore found in larger numbers in the Strait of Juan de Fuca, the San Juan Islands, and Puget Sound than near the outer coast. However, a few animals do occur, from Grays Harbor north along the continental shelf.
(7) Fin whales are a pelagic species, occurring in Washington as a part of their summer range. They winter in tropical waters. The population of fin whales was originally second only to gray whales off the outer Washington coast. They occur along the coast north of Grays Harbor, but their major area of concentration is in the Cape Flattery region and the mouth of the Strait of Juan de Fuca. Their food consists primarily of euphasiids, and they travel in small pods of two to five individuals.
(8) Sei whales, like fin whales, occur in Washington as part of their summer range, wintering to the south of California. This species is more restricted to the outer coast, occurring from Grays Harbor northward along the continental shelf. They travel in small pods of two to five individuals.
(9) Humpback whales also occur in Washington as part of their summer range, making extensive seasonal migrations to wintering grounds south of California. They migrate in close proximity to the coastline, and are often visible from shore. They travel individually or in groups containing up to 25 animals. They are found along the outer coast north of Grays Harbor, and in most of the inside waters (although rarely in Puget Sound). They feed on euphasiids and small schooling fishes (Everitt et al. 1980).
Resident Sea Mammals of the Outer Coast
For all the preceding sea mammal species, the critical factor for prehistoric human exploitation was the proximity to annual migration routes. For other resident species, however, the critical factor for human exploitation was the proximity of haulouts and/or rookeries. These resident species include the harbor seal (Phoca vitulina), the sea-otter (Enhydra lutris), and various species of toothed whales such as dolphins, porpoises, and orcas.
(1) Harbor seals are distributed in all of the coastal waters of Washington where they reside on a year-round basis. For protection from predators, they haul out on exposed rocks or sand bars. Since shellfish form a primary food, they frequent tidal flats and shallow bays in order to obtain them. Because salmon are another important food, harbor seals are also found near the mouths of salmon streams during spawning migrations and are sometimes known to follow salmon considerable distances upriver. Greatest concentrations of seals are found to the south of the study area, near Grays Harbor and Willapa Bay (Wahl 1977). To the north, at least four major haulouts are located from Toleak Point to north of the Hoh River (Newby 1973). The northernmost area of the outer coast contains fewer haulouts for harbor seals, particularly in comparison with areas further east along the Strait of Juan de Fuca (Johnson and Jeffries 1977; Sanger 1965).
(2) Sea-otters originally occurred as resident species along the outer coast, from the Ozette area south to the Columbia River (Scheffer 1940). Their preferred habitat includes exposed rocky islands that offer shelter from storms, and underwater reefs and large kelp beds for obtaining their preferred foods—various bottom invertebrates in the subtidal zone (e.g. sea urchins). Along the outer coast, major occupied areas include the Ozette area, Cape Johnson, James Island, the LaPush region, Destruction Island, and Point Grenville. Their low mobility and low reproductive rate make them easily subject to overexploitation.
(3) Toothed whales are distributed throughout the outer and inside waters of Washington, and are generally present on a year-round basis. Seventeen species are present, many of which were used by prehistoric people, although not as consistently as the larger baleen whales or the seals and sea-otters which haul out in nearshore locations. These species include the following: Pacific whiteside dolphin (Lagenorhynchus obliquidens), whitehead grampus (Grampus griseus), Orca or killer whale (Orcinus orca), Harbor Porpoise (Phocoena phocoena), Dall's porpoises (Phocoenoides dalli), bottlenosed whale (Berardius bairdii), and goosebeak whale (Ziphius cavirostris), as well as a number of rarer or incidental species.
Sea-mammals of the Inner Coast
Sea-mammal taxa become depauperate as one moves from west to east along the inner coast, and this is a trend that is repeated with sea-birds to be discussed below. Sea-otters are essentially absent from the region, although river otters (Lutra canadensis) increase in numbers, particularly to the east of the Elwha River (Everitt et al. 1980). Fur seals are also essentially absent from the region, except for the outer Juan de Fuca Strait. Limited numbers of fur seals, mostly yearlings, venture into the outer Strait during annual migrations, with "most sightings occurring from January through March" (Everitt et al. 1980:28). The California sea lion occurs regularly in the outer portions of the Strait of Juan de Fuca, having at least 11 haulouts in that region on both sides of the Strait; however, they occur rarely east of Race Rocks on Vancouver Island, opposite the mouth of the Elwha River (Everitt et al. 1980:19). Northern sea lions are more widely distributed in the region, although in fewer numbers (a current maximum population of 200 as opposed to ca. 300 California Sea Lions). At least ten haulouts are known for the area, ranging from Tatoosh Island on the west to Dungeness Spit on the east. No haulouts are known for the southern Puget Sound or Hood Canal region, however, and relatively fewer animals penetrate into this area. As on the outer coast, both species of sea-lions are absent in summer (June through August or September), with peak densities from October through April (or May). Northern elephant seals do not haul out in the region, but are widely distributed in all regions except for Hood Canal (Everitt et al. 1980:47). Largest numbers of sightings of these pelagic seals occur in the area between the Elwha River and the mouth of the Hood Canal.
By far the most widely distributed and densely populated seal in the region is the harbor seal. This species is a permanent resident in the Strait of Juan de Fuca and Puget Sound. The number of harbor seals known for this region is approximately the same as the population of the northern outer coast, ca. 1,200-1,300 animals in each case (Johnson and Jeffries 1977; Everitt et al. 1980). (These populations are, of course, dwarfed by the much larger numbers of animals inhabiting the San Juan Islands and the southern outer coast). Of the inner coastal populations, ca. 100 are located in the outer Strait, ca. 350 in the inner Strait and northwestern Puget Sound, and ca. 725 in the Hood Canal region (Everitt et al. 1980). Six major haulouts for these animals are known for the outer Strait region and five haulouts for the inner Strait and northwestern Puget Sound (see Figure 3.5; Everitt et al. 1980:37). Approximately 280 seals are known to haul out in the outer Strait, whereas as many as five times that number may haul out in the inner Strait. In general, therefore, numbers of available harbor seals increase from west to east. To this is added the behavioral feature that harbor seals in the inner Strait and Puget Sound waters are more likely to haul out at high as well as low tides; the reason for this is probably related to the lower tide in the region and the availability of sandy haulout areas at high tides. For all of these reasons, more intense exploitation of harbor seals would be expected in the eastern portion of the region.
Among the baleen whales, only two are currently found in any numbers along the inner coast: the gray whale and the minke whale. Of these, only the gray whale is distributed throughout the region in any numbers, being found in all seasons as far east as the mouth of the Hood Canal. Minke whales are largely restricted to the San Juan Island region, although some pods are known to utilize the area between Port Angeles and Port Townsend during the spring and summer (Everitt et al. 1980:52). Formerly, however the humpback whale was also widely distributed throughout the inner coast, including Puget Sound. Other species, such as fin whales, are strictly pelagic and do not enter the waters of the inner coast.
Among the toothed whales, only three achieve significant populations in the inner coastal region: the Orca (Orcinus orca), the harbor porpoise (Phocoena phocoena), and the Dall porpoise (Phocoenoides dalli). Of these, the harbor porpoise is generally restricted to the San Juan Island area, occasionally utilizing the region between Port Angeles and Port Townsend (Everitt et al. 1980:61). Orcas are probably somewhat more abundant in the region, with up to several hundred individuals present in summer, when populations are greatest (Bigg and Wolman 1975). Dall's porpoise is probably the most abundant in the area at all seasons of the year, with ca. 300 permanent residents (Everitt et al. 1980:62). They have an interesting seasonal round, however, utilizing the outer Strait of Juan de Fuca almost exclusively during the fall and winter and the inner Strait primarily in winter and spring (Everitt et al. 1980:63). Other species of toothed whales are almost exclusively pelagic and do not use the inner coastal waters.
Ethnographic Information on Sea-Mammal Hunting
Ethnographic evidence on sea-mammal use is perhaps greater than that for any other resource, particularly in terms of technology. Swan (1869) noted the use of several species of sea-mammals by the Makah, including the northern sea lion, northern fur seal, harbor seal, sea-otter, California gray whale, fine whale, right whale, blue whale, sperm whale, and a number of toothed whales, including porpoise, orcas, and pilot whales. Among the whales, however, Swan (1869) clearly states that only the gray whale was used consistently, right whales were occasionally hunted but were more likely to be retrieved as beached or stranded individuals. It is interesting, however, that no mention is made of the humpback whale, since archaeological data suggest that this species was quite important (Huelsbeck 1983b).
In contrast to the Makah pattern, ethnographies for groups to the south on the outer coast show much less focus on whaling. The Quileute engaged in a limited amount of whaling, including both the gray and fin whales (Collins 1892; Pettitt 1950), although it has been suggested that this was a relatively late phenomena resulting at least partially from contact with the Makah to the north (Taylor and Bosch 1980; Fleisher 1984). Still further south, the Quinault Indians appear to have undertaken whaling in a much more systematic fashion, taking large whales as much as 12-30 mi offshore (Olson 1936). The Queets division of the Quinault, however, were apparently not whalers but they, along with the Quinault made extensive use of stranded whales (Olson 1936). To the south of the Quinault, almost no use was made of whales except for those that were occasionally stranded. Fur seals, sea-lions, harbor seals, sea-otters, and porpoises, however, were all important to the groups to the south of Ozette (Pettitt 1950; Olson 1936). However, since the Quinault had to venture from 10 to 25 miles offshore to obtain fur seals, they were emphasized less in this area than to the north. Correspondingly, all of the other species, particularly the sea-lion, were emphasized more.
Among the Klallam, whale hunting was carried on as an active pursuit (unlike the Indians of the southern outer coast), but were only pursued when spotted from shore (Gunther 1927:204). Sealing probably was more important economically than the pursuit of other sea mammals and involved complex technology, ceremony, and sharing rules. Hunting for porpoise and "blackfish" (toothed whales) appears to have been a secondary pursuit, possibly because these animals were neither large in size (as the baleen whales) nor easy to pursue at haulouts (as the harbor seals). Among the Twana of the Hood Canal region, whales were not hunted but beached whales were welcomed and utilized (Elmendorf 1960). Harbor Seals were clearly the most important species, being trapped, netted and harpooned; porpoises were secondary in importance, while sea-lions were only occasionally utilized when they strayed into the Canal area.
Salmonid Resources
This class of resources is sufficiently important to warrant more detailed consideration than most of the marine fishes. Also, anadromous fish do not neatly fit into the category of marine resources; even though they achieve much of their body weight feeding on marine pastures, they spend important phases of their lives in freshwater rivers and lakes and are available at considerable distances from the sea.
Anadromous fish species native to the Olympic National Park include chinook salmon (Oncorhynchus tshawytscha), coho salmon (O. kisutch, sockeye salmon (O. nerka), chum salmon (O. keta), and pink salmon (O. gorbuscha), steelhead trout (Salmo gairdneri), cutthroat trout (Salmo clarki), and Dolly Varden (Salvelinus malma). Although there are substantial variations in the seasons of migration even within individual species, the general seasonal timing of the fish runs is illustrated in Figure 3.6.

|
| Figure 3.6 Generalized schedule of salmon spawning migrations for the Olympic Peninsula (from Phinney and Bucknell 1975). (click on image for a PDF version) |
Anadromous fish runs on the Olympic Peninsula probably represent some of the best preserved wild populations remaining in the Northwest primarily due to the protection of habitat that has been afforded by the Olympic National Park. This is not to suggest, however, that the runs in the rivers of the Peninsula have not suffered significantly from a number of factors (Brown 1983). Although we do not assume that the recent patterns in these resources directly reflect conditions at all times in the past, these historical data do provide the best available information for modeling fish resources during the past several thousand years.
River basin morphology is a major variable conditioning the nature of fish runs in any region and this is also the case on the Peninsula. Due to the radial drainage structure of river valleys surrounding the central Olympic massif, all the rivers are relatively short and have steep gradients in their upper portions. Insofar as fish production is correlated with the availability of freshwater spawning habitat, the small size of these drainages places a low upper limit on the quantity of fish that can be intercepted in a particular river.
An important aspect of drainage morphology that influences fish distribution within river systems is the presence of impassable falls and cascades. These have been mapped in considerable detail by the Washington Department of Fisheries (Phinney and Bucknell 1975; Williams et al 1975) and this information was used in mapping the maximum distances salmon travel upstream in the major valleys of the Peninsula. There are two interesting points regarding patterning in the distribution of these obstructions on the Peninsula. The first is that alpine glaciers carved rounded valley bottoms in the major river valleys (Hoh, Queets, Quinault) but left many tributary streams entering these as hanging valleys. Most streams in these tributary valleys enter the trunk drainage over cascades or falls that are impassable to migratory fish. The second point is that the streams draining the eastern Olympics (excluding the Skokomish) are much steeper in gradient than elsewhere on the Peninsula. The major consequence of this fact is that fish migrations are blocked in most east side drainages no more than a few miles above saltwater and in most cases well outside the boundaries of the Park. Figure 3.7 shows the extent to which salmon runs potentially penetrate the Olympic National Park's central section. The Elwha, however, is a moderate gradient valley along its main stem and prior to the construction of the Elwha Dam in 1910, fish probably moved considerable distances into the Park in this river.
|
| Figure 3.7 Accessible spawning habitat for anadromous fish in the major drainages of the Olympic Peninsula (based on Phinney and Bucknell 1975). (click on image for a PDF version) |
From the perspective of latitudinal gradients in anadromous fish distributions in Western North America, the Olympic Peninsula falls within the central zone where both northerly and southerly species overlap in their distribution (Schalk 1977). On the one hand, pink, chum, and sockeye, represent the dominant species in the more northerly areas of the continent. On the other hand, the chinook, coho, and steelhead tend to dominate in more southerly rivers. Although the particular mix of species and runs of salmon in a given river depends largely on the amount and diversity of spawning habitat present, all five species of Pacific salmon are well represented in the rivers of the Peninsula. Mild winters associated with the exceptional maritime climate must partially account for diversity of salmon species in this region. Similarly, the presence of extensive glaciers and snowfields in the mountains contributes to stream flow during the dry summers when low run-off is typically a limiting factor for fish production. Compared to most other coastal drainages of Oregon and Washington, those of the Olympics are exceptional in this respect because they draw tribute from higher elevations with extensive snowfields.
Some rivers of the Peninsula have high species diversity as well as run diversity. There are, for example, spring, summer, and fall runs of chinook and early and late chum runs in certain rivers. Table 3.2 provides a list of the twelve largest rivers of the Peninsula and the runs of salmon documented historically for each. A number of points illustrated in this table are worthy of comment. Particular attention will be afforded the geographical patterns in species of salmon around the Peninsula.
Table 3.2 Salmon runs in the twelve largest rivers of the Olympic Peninsula (data from Phinney and Bucknell 1975).
| River | Spring Chinook |
Summer Chinook | Fall Chinook |
Coho | Pink | Sockeye | Early Chum | Late Chum |
Total Runs |
| Ozette | X | X | X | X | 4 | ||||
| Quillayute | X | X | X | X | X | X | X | 7 | |
| Hoh | X | X | X | X | X | X | 6 | ||
| Queets | X | X | X | X | X | X | 6 | ||
| Quinault | X | X | X | X | X | X | X | 7 | |
| Skokomish | X | X | X | X | X | X | X | 7 | |
| Hamma Hamma | X | X | X | X | X | 5 | |||
| Duckabush | X | X | X | X | X | X | X | 7 | |
| Dosewallips | X | X | X | X | X | X | X | 7 | |
| Quilcene | X | X | X | X | X | 5 | |||
| Dungeness | X | X | X | X* | X | 6 | |||
| Elwha | X | X | X | X | X | 5 | |||
The larger rivers of the Peninsula support at least four of the five species of Pacific salmon or between four and seven distinguishable runs of these species. The sockeye salmon, due to their usual preference for drainages with lake systems, occur in significant numbers only in the Quinault and Ozette rivers. Lake Crescent apparently was inaccessible due to natural obstructions in the lower reach of the Lyre river (Phinney and Bucknell 1975) and, therefore, not a sockeye lake. Spring and summer chinook require drainages with substantial dry season run-off and it is noteworthy that these runs are not as well represented in the shorter drainages that originate on the dry side of the mountains (e.g., Quilcene, Hamma Hamma) or which drain relatively low foothills that accumulate minimal winter snowpack (e.g., Ozette). Smaller rivers of the Peninsula which do not have their sources deep in the mountains usually support runs of fish that make their spawning migration during the fall after the rains begin (i.e. coho, chum, pinks, and fall chinook). Chum salmon, at least in small numbers, are ubiquitous in the larger rivers of the Peninsula but the early chum runs appear to be absent from west slope river valleys. [2] Coho are undoubtedly the most widespread of the salmon species and occur in virtually all streams and creeks of the region. Pink salmon are decidedly uneven in their distribution throughout the Peninsula. Small numbers occur in both even and odd years along the west slope of the Peninsula. It is probably significant that these numbers are so small that some studies of salmon spawning areas indicate no pink salmon west of the Elwha on the Peninsula (Atkinson et al. 1967:88). In Hood Canal Rivers, in contrast, pinks are not only the most abundant of all salmon species but only occur in significant numbers during odd years. The Elwha and Dungeness parallel the Hood Canal drainages having pink as the dominant salmon species, but at least in the case of the Elwha, this may be a relatively recent post-reservoir development.
As a rough way of illustrating major east side and west side contrasts in salmon species abundances, historic escapements (Phinney and Bucknell 1975) have been compared for two areas from which comparable data were available (see Figure 3.8). Numbers of individuals in average runs for each species were converted to percentages of the total salmon biomass (in lbs) entering rivers of the Hood Canal Basin and the Soleduck-Hoh Basin. There are clearly dramatic differences in relative abundance.
|
| Figure 3.8 Escapement (by weight) of various salmon species in the Hood Canal Basin versus the Soleduck-Hoh Basin (based on estimates for 1966-1971 from Phinney and Bucknell 1975). (click on image for a PDF version) |
East slope rivers tend to be dominated quantitatively by pink salmon with chums of secondary importance. [3] Chinook and coho strongly dominate the fishery in the Soleduck-Hoh basin streams with sockeye in a tertiary position. There is no reason to doubt that this broad difference probably has long standing significance for rivers of the western Peninsula though sockeye would have been more important in the Quinault River.
It is estimated that the entire Hood Canal Basin has a total of 70 miles of river valleys for migration, spawning, and rearing of anadromous fish (Williams et al. 1975). This compares to individual west slope rivers such as the Quillayute with 374 miles of accessible river habitat and to the Quinault with 377 miles (Phinney and Bucknell 1975). Pink and chum, in contrast to chinook, coho, and sockeye, do not typically migrate any great distance up the rivers of North America. In general, the scarcity of accessible salmon habitat in the east slope river valleys, would have always favored those species such as pink and chum which spawn near saltwater and do not require freshwater rearing areas. One consequence of interest here ethnographically and archaeologically would be that the salmon fishing season could extend for a considerably longer portion of the year on the west side of the Peninsula than on the east side where most of the salmon biomass in most streams had to come from fall spawning migrations. The Skokomish River, due to its size and distinctive character, is exceptional for having anadromous fish runs through a greater portion of the year than other streams that enter Hood Canal.
To summarize, there is much greater variability in the salmon resources around the Peninsula than is frequently obvious from archaeological or ethnographic literature of this region. The greatest contrast within the Peninsula region is that between the east and west slope rivers of the Olympics. East side rivers have drastically smaller areas of habitat for the migration, spawning and rearing of salmonids. Salmon are all but excluded from entry into the Olympic National Park from the rivers east of the Elwha. West side rivers like the Quillayute, Hoh, Queets, and Quinault are characterized by extensive areas of accessible salmonid habitat and larger, more diverse runs of these fish. Correlated with these major east-west contrasts there are significant differences in the relative importance of the five species of salmon. West side rivers are dominated by chinook, coho and sockeye runs; east side by pink and chum runs. East slope salmon appear to be more dispersed spatially and more seasonally limited to the fall than are those of the west slope rivers. The minor exceptions to this overall pattern involve the Elwha, the Skokomish, and the small drainages of the northwestern corner of the Peninsula (Ozette, Sooes, Hoko, and Psysht). The salmonid resources of the Elwha may have been intermediate in terms of species proportions to the east and west side rivers characterized above. In terms of the drainage size and structure and habitat area, however, the Elwha compares most closely to west side rivers. Similarly, the Skokomish is larger, supports much more extensive salmon habitat, and contains more substantial moderate gradient lower reach than other Hood Canal rivers. The rivers of the northwestern corner of the Peninsula are notable in the low relief areas which they drain. As a result of this characteristic, they are small rivers with markedly seasonal flow regimes and low summer flows. Salmon runs in this area, with the exception of the Ozette River sockeye, were very limited.
These points are of importance because of the common tendency to generalize the results of archaeological research in better known areas to the surrounding regions. The focus of archaeological research in this region has been disproportionately located in the northwestern Peninsula and the structure of resources in that area is sufficiently different from the rest of the Peninsula that it would be unlikely that the archaeological patterning there is in any sense typical.
Finally, the distribution of anadromous fish around the Peninsula seems to account for a significant amount of variability in ethnographic settlement systems. It is clear that the rivers with large and diverse runs of salmon such as the Quinault, Queets, Quillayute, Elwha, and Skokomish supported winter village communities for many miles inland. In all other areas of the Peninsula, salmon resources were apparently exploited by people whose winter villages were based exclusively on saltwater.
Marine Fish of the Outer Coast
Excluding the anadromous fish species, there are essentially two classes of marine fish species available to inhabitants of the Outer Coastal zone: nearshore and offshore. Nearshore fish are generally surface feeders and most are available throughout the year although a few may be present only seasonally, as during the breeding season (e.g. herring). Offshore fish are generally deeper-water, non-schooling species that must be obtained with the use of boats.
Shallow water nearshore species include the following:
1) sharks, especially the spiny dogfish (Squalus acanthias);
2) skates (Raja spp.)
3) herring (Clupea harengus)
4) walleye pollock (Theragra chalcogramma)
5) perch (embiotocidae; 5 genera)
6) prickleback (2 genera)
7) gennel (2 genera)
8) rockfish (Sebastes spp.)
9) sablefish (Anoplopoma fimbria)
10) greenling (Hexagrammos spp.)
11) cabezon (Scorpaenichthys mormoratus)
12) sculpin (Cattidae; 16 genera)
13) poacher (7 genera)
14) snailfish (Liparis spp.)
15) sanddab (Citharichthys spp.)
16) smelt (Hypomesus pretiosus)
Deeper water offshore species include:
(1) Pacific cod (Gadus macrocephalus)
(2) tomcod (Microgadus proximus)
(3) lingcod (Ophiodon elongatus)
(4) sole (7 genera)
(5) halibut (Hippoglossus stenolepis)
(6) flounder (Platichthys stellatus)
(7) turbot (Reinhardtius hippoglossoides)
(8) flatfish (Plaeuronectiformes)
Miller et al. (1980) and Cross (1981) provide fuller accounts of the ecology and distribution of these species.
Marine Fish on the Inner Coast
In addition to the contrast between the inner and outer coasts in terms of the availability of whales, sea-otters, and other sea-mammals, there is a notable contrast in the availability of marine fish. In the inner coastal region, offshore species such as halibut and lingcod are generally less available, while nearshore species are often found in greater numbers as a result of the relatively shallower, quieter coastal zone. In this zone, studies by beach seine and tow-net (Miller et al. 1980) have demonstrated that the following fishes were most abundant in the nearshore environment: herring; sculpin; sole; flounder, tomcod; perch; dogfish; and skate.
Ethnographic Accounts of Marine Fish Use
Ethnographic accounts of fish use by the Makah (Swan 1869) indicate that at least eight species of fish were economically important. Among these, clearly the most important was halibut, followed by salmon and lingcod. Somewhat less important were a variety of nearshore species, including sculpins and "groupers" (possibly rockfish and/or greenlings). Following this was tomcod, while true cod was "taken in limited quantities," owing to "the extreme depth and swift currents of [Juan de] Fuca Strait" which made it difficult to fish in "thirty to forty fathoms of water" (Swan 1869). Perch and herring are also mentioned, although primarily as bait. Finally, dogfish was noted as a resource, primarily for the liver oil prepared from it and used for trade.
From the Quileute south, the deep-water, offshore species (halibut and lingcod) become secondary resources, while the small, nearshore schooling species decline even further in importance. Instead, the anadromous fish, particularly salmon, but also trout, smelt, and eulachon or candlefish became first in importance. This is seemingly due to the size and nature of the river systems found in both the Quileute and especially Quinault areas, and to the lack of emphasis on an offshore hunting technology. Among the Quileute, salmon were most important, followed by smelt and herring [4] (Pettitt 1950:5). Among the Quinault, the anadromous fish were followed in importance by halibut, lingcod. sole, flounder, and the sea bass (Ciynoscion nobilis), of which the latter is incidental in the Ozette area to the north (Friedman 1976). Mentioned last again were the herring and other nearshore fish.
Among the Klallam, fishing for halibut, lingcod, and flounder was still relatively important, secondary only to fishing for salmon and steelhead trout. These species were obtained by line fishing (halibut, lingcod) or spearing (flounder) between April and September (Gunther 1927:199). Halibut, in particular, were more likely to be taken off fishing banks between Jamestown and Port Angeles (Gunther 1927:201). Fishing for nearshore species, particularly herring, and smaller anadromous species (smelt and candlefish) was less important than the deep water fishing. By far the most important was fishing for four species of salmon and steelhead trout, using a wide variety of methods including trapping, trolling, gill-netting, spearing, and linefishing in the rivers.
Again, for the Twana to the east, fishing for salmon and steelhead trout was most important, but these species were taken with traps, dipnets, and harpoons rather than by trolling or gillnetting. Second in importance were herring, which were impounded in tidal-shore enclosures and net traps (Elmendorf 1960), various other nearshore species such as skate and tomcod, and bottom fish like halibut were occasionally taken, but only in the northern part of the Hood Canal (Elmendorf 1960:57). Sturgeon were also found to a minor extent in this area, and along with smelt were less important anadromous species that were used for food.
Seabirds of the Outer Coast
Besides supporting populations of shellfish and other invertebrates, the nutrient base of the outer coast of the Peninsula helped to support substantial sea-bird populations These may be subdivided into two groups: offshore and nearshore species.
Important offshore species include the loons (Gavia spp.), albatrosses (Diomedea spp), grebes (Podiceps spp.), shearwaters or "muttonbirds" (Puffinus spp.), the Northern Fulmar (Fulmarus glacialis), cormorants (Phalacrocorax spp.), Great Blue Heron (Ardea herodias), black oystercatcher (Haematopus bachmani), sea gulls (Larus spp.), black-legged Kittiwake (Rissa tridactyla), common tern (Sterna hirunda), common murre (Uria aalge), pigeon guillemot (Cepphus columba), tufted puffin (Lunda cirrhata), and a variety of smaller alcids (Sanger 1970; Wahl et al. 1981; Salo 1975). Certain areas, such as Tatoosh Island near Cape Flattery, harbor major breeding colonies, particularly for gulls and alcids. In fact, sea-bird colonies are larger in rookeries, especially island rookeries, along the outer coast than for any other area of the Peninsula region except for Protection Island near Port Townsend (Wahl et al. 1981). Birds that breed on these islands also tend to maintain colonies during the rest of the year. Within the outer coastal region, furthermore, there is a change in distribution of sea-birds that parallels the distribution of invertebrate resources. North of the Hoh River region, the abundance and diversity of sea-birds is significantly greater because the offshore islands associated with the complex, rocky coastline offer greater opportunities both for bird nesting and protection from predators (c.f. Figure 3.9). Large sea-bird colonies tend to be continuously distributed throughout this region, except for Mukkaw Bay between Cape Flattery and Cape Alava.

|
| Figure 3.9 Sea bird rookeries of the northwestern Olympic Peninsula (after Proctor et al. 1980). (click on image for a PDF version) |
The nearshore species found on the western Olympic coast are dominated by waterfowl (swans, geese, and ducks); these are prevalent on both the outer and inner coasts of the Peninsula. On the outer coast, these species utilize microhabitats different from those used inland: islands, small bays, tidal inlets, coastal meadows, shallow margins of salt marshes, sloughs, and even tidal flats (Salo 1975; Brittell et al. 1976). One high density zone for waterfowl is in Mukkaw Bay between Cape Flattery and Cape Alava, where appropriate habitat exists. Some other waterfowl species, however (e.g. the Harlequin duck, (Histrionicus histrionicus) will utilize rocky seacoast areas, sharing them with the sea-birds. Still other species (e.g. the hooded merganser, Merous serrator) use areas adjacent to lakes such as Ozette Lake. In addition to waterfowl, other nearshore species on the outer coast include large shorebirds such as the long-billed curlew, wandering tattler, Lapland Longspur, Marlin, and Turnstones (Salo 1975; Brittell et al. 1976).
Some of these species (Harlequin duck, black oystercatcher, glaucous-winged gull, tufted puffin, smaller alcids) are year-round residents, found in nearly equal numbers at various times of year. Others vary in number seasonally. Overall density peaks for the outer coast appear to be greatest in spring, with a second peak in fall; densities at those times may exceed 1,300 birds/km2 (Wahl et al. 1981). Numerous species are also winter residents on the outer coast, but may feed offshore or breed in other areas during the summer period.
Sea-birds of the Inner Coast
Habitats for sea-birds differ substantially on the inner and outer coasts. The inner coastal regions harbor few rocky, offshore islets where nesting sea-birds are safe from predators. Resources for many offshore, pelagic-feeding birds are not present. On the other hand, the quieter lagoonal environments in many areas (particularly east of the Elwha River) offer excellent habitat for waterfowl, including many migratory species. These trends are reflected in the species observed along the inner coast. In the outer Juan de Fuca Strait area, for example, some pelagic sea-birds such as albatrosses, shearwaters, and fulmars, are absent; others such as petrels, cormorants, gulls, murres, puffins, guillemots, and other alcids are found in significant numbers. In the inner Juan de Fuca Strait/northwest Puget Sound region, however, very few of these birds are found, except for some cormorants and guillemots (Wahl et al. 1981). The one exception to this pattern is on Protection Island, where large numbers of gulls and rhinoceros auklets are found. Although the offshore sea-bird fauna becomes increasingly depauperate as one moves from west to east, waterfowl increase in numbers and densities because of increase in available habitat.
Ethnographic Information on Sea-bird Use
Ethnographic accounts on the use of avifauna discuss the use of both sea-birds and waterfowl. Swan (1869:25) notes that the Makah were very fond of sea-birds, which were
taken in great numbers on foggy nights, by means of spears. A fire of pitchwood is built on a platform at one end of the canoe, and by the glare of its light, which seems to attract the birds, the Indian is enabled to get into the midst of the flock, and spear them at his leisure.... These, after, being picked, and very superficially cleaned, are thrown promiscuously into a kettle, and boiled and served as a feast.
Swan also records the following species that were used by the Makah: loons, grebes, cormorants, herons, eagles, cranes, sandpipers, sea gulls, guillemots, and various species of "divers" (alcids) and waterfowl. A few terrestrial species were apparently used, including grouse, crows, kingfishers, and woodpeckers. For the Quinault, Olson (1936) also notes utilization of loons, cormorants, herons, eagles, sea gulls, and various waterfowl, and also cites the use of some terrestrial forms such as grouse. Gunther (1936) reiterates essentially the same list of species in her report on the "Zoological Knowledge of the Makah". Unfortunately however, no attempts have been made to rank order the ethnographic use of these species. Gunther (1927) notes that duck hunting was the primary bird subsistence pursuit among the Klallam, primarily in the region between Port Angeles and Port Townsend; swans and geese were also important, but only gulls and eagles were occasionally eaten. Elmendorf (1960) also emphasizes waterfowl hunting for the Twana, including "geese, brant, and several species of duck."
Discussion
This chapter has focused upon a characterization of the historic and modern ecosystems of the Olympic Peninsula. Although this characterization would be questionable to project into the distant past, the recent environment provides the most useful frame of reference for the consideration of how past environments might have differed. Chapter 5 examines regional paleoenvironments against the frame of reference provided by this chapter.
The Olympic Peninsula is a region of exceptional environmental diversity in both its terrestrial and marine ecosystems. Along the entire Northwest Coast, there is probably not another area of comparable size that is as internally varied. This intraregional environmental differentiation inevitably contributes to an equivalent degree of diversity in the nature of human land use systems in the past.
In general, the coniferous rainforests dominate the region and these forests have very high primary productivity. From the viewpoint of human subsistence, however, the terrestrial resources in this ecosystem are rather poor. Edible vegetal resources are strongly associated with the small openings or prairies in the nearly continuous forest but the historical evidence suggests that these openings are both small and infrequent. Exploitable secondary biomass in a high primary biomass environment is typically unproductive and the Olympic Peninsula conforms to this general pattern. Ungulate densities in this environment do not compare favorably with drier, more open settings. In addition to relatively low productivity for large animals, species diversity is also quite low and this is at least partially due to the "island-like" character of a mountain massif that is separated from other ranges by extensive lowlands.
The diversity and abundance of marine resources varies enormously moving around the Peninsula. There is a progression from the open coast of the western Peninsula through the broad Straits of Juan de Fuca and the protected fiord of Hood Canal. In the next chapter, these differences are considered from the perspective of the ethnographic land use systems around the Peninsula.
ENDNOTES
1 Gunther (1927:272) mentions a prairie on the upper Elwha River but gives no other locational details. A prairie is also mentioned by Elmendorf (1960:36) in the area occupied by the Vance Creek band of the Twana—a group he describes as "wholly inland in habitat." Still other prairies were located south of the Skokomish river and Twana groups gathered camas at these places (Elmendorf 1960:126,128).
2 Another factor implied by the occurrence of early chum runs in the east side rivers is that water temperatures drop relatively early in the fall there because these streams do not flow through a coastal lowland.
3 According to Elmendorf (1960:61), however, chums "provided the majority of fish taken in the Skokomish in aboriginal times."
4 Pettitt's (1950:5) reference to herring is ambiguous insofar as he may have been using the term to describe smelt.
| <<< Previous | <<< Contents>>> | Next >>> |
schalk/chap3.htm
Last Updated: 16-Nov-2009