|
NATIONAL PARK SERVICE
Fauna of the National Parks of the United States No. 7 The Wolves of Isle Royale |

|
RESULTS—THE TIMBER WOLF AND ITS ECOLOGY
NO WOLF or wolf carcass was handled during this investigation, so no vital statistics are available for the Isle Royale animals. However, these wolves undoubtedly are of the same subspecies as those on the nearby mainland—Canis lupus lycaon. Stenlund (1955) examined approximately 150 wolves of this subspecies from northern Minnesota and found that the body length of most males was 43 to 48 inches; the majority of females measured 41 to 45 inches. Females averaged about 61 pounds, and males about 78, although some males weighed over 100 pounds. Observations of most Isle Royale wolves from about 50 feet, and of one from 15 feet (figure 30), indicate that these animals are about the same size as those from Minnesota. All of the park's wolves are gray, with so little color variation that individuals are indistinguishable on this basis.

Figure 30—An Isle Royale timber wolf at 15 feet.
In most of the wolf's range, airplanes are used to hunt the animals. Minnesota wolf-hunters claim that wolves have learned to vacate open areas at the sound of an airplane, and Stenlund (1955) and Cole (1957) provide evidence for this contention. On Isle Royale, Cole found that the wolves reacted unpredictably to his light craft. At times they bolted for the nearest cover, but in other instances they calmly watched the plane pass several times within 100 feet.
The first time we encountered wolves they (six) showed little concern until the plane approached to within about 200 feet; then they arose from their beds (figure 31).

Figure 31—Attitude of wolves upon the initial approach of our
aircraft.
Each time we passed within 100 feet, they rushed toward the craft. This continued until we left. When the plane drew near nine other wolves on a ridge a few miles away, they ran onto the ice and tried to chase the craft. A few hours later, the groups were together and responded to us as they had earlier. These wolves were seen again 2 days later; 200 feet below us, they showed little concern.
Throughout the rest of the first winter study period they remained oblivious to our presence, except once when continually buzzed at 75 feet. (Unless special conditions warranted, we usually flew at about 300 feet.)
This large pack apparently remained conditioned to the aircraft for a year, since low passes during the first day of the 1960 study period failed to disturb them. Even when we landed within 60 yards of the animals they stood their ground. Several ran back and forth on the ice for 1-1/2 minutes and started toward the plane a few times, but they all finally ran into the woods and howled. When relocated the next day, they were unafraid. Through out this study period the wolves appeared unconcerned at our presence (figure 32). Even when buzzed 10 times at about 40 feet on the last day, they merely stood around and watched.

Figure 32—The unconcern shown the aircraft by the large pack
throughout most of the study.
Our first approach in 1961 frightened a few of the wolves in this large pack, but most remained unafraid. This disparity in behavior between individuals was noticed throughout February and March 1961, and probably resulted primarily from differences in social arrangement of the pack and the corresponding variations in social status of individuals.
Packs encountered less frequently showed more concern about the airplane. Apparently, wolves become conditioned by the continued presence of an aircraft that causes them no harm. Burkholder (1959) in Alaska found that wolves which were at first afraid of his craft eventually became accustomed to it.
Several times when the Isle Royale wolves were encountered while resting, they became aroused and began traveling a few minutes after the plane approached. They did not seem to be unduly concerned over the aircraft but may have been bothered by the noise. However, this was not considered enough of a disturbance of natural activity to complicate the results of our observations.
Wolves in most areas are known to be afraid of man, and experiences with Isle Royale wolves demonstrate the extreme to which this is true. On three occasions I chased 15 wolves from a moose carcass upon which they had just begun to feed. Although a few individuals were reluctant to leave until I approached to within, in one case, about 40 feet, all finally retreated and failed to return until several hours after the carcass had been examined. The manner in which the wolves left one carcass was especially interesting. When I approached to within 150 yards, most of the pack ran. Six animals continued tugging at the carcass until I got to within about 60 feet, and then two looked up at me and unceremoniously left. The other four, heads buried in the carcass, apparently received no signals from these individuals. They didn't detect me until I was about 40 feet away. Suddenly all jumped up and ran about 75 yards, stopped, looked back, and then continued to the rest of the pack, about 150 yards away.
On one occasion, after I had disturbed the wolves and examined their kill, part of the pack made a new kill while the others returned and fed on the old. The former animals apparently did not return to the original kill for about 2 days. The behavior of individuals during several other close-up encounters during winter and summer attested to the Isle Royale wolves' fear of man. A pack of three even were afraid of the human scent on a package of crackers tossed from the aircraft; each took one sniff and dashed off. However, the wolves were completely unafraid of docks, cabins, and other manmade structures which had no recent human scent.
Wolf Numbers
 Figure 33—Wolf tracks in snow. |
The composition of the Isle Royale wolf population makes possible a reasonably precise count but renders difficult an absolutely complete census. The main pack, containing 15 to 16 members, usually was relatively easy to locate. However, groups of three and two, and lone individuals, also were sighted. (These smaller groups were difficult to find and keep track of.) Thus, censusing involved finding and counting the large pack and then trying to locate all other groups. The wolves' preference for traveling along the Isle Royale shore or on lakes was most important in the success of the censuses. Censuses were attempted on the first or second day after a fresh snowfall, or under the following combination of circumstances: (1) the known whereabouts of the large pack, and (2) the discovery of a recent kill made by another pack (thus often allowing the prompt locating of this pack), and/or (3) the accidental sighting of other wolves. During censuses, we flew at 300 to 500 feet altitude and surveyed the entire shoreline and all major lakes for wolves or tracks (figure 33). Tracks were followed until the wolves were found, if possible. Undoubtedly, no large packs escaped detection, but perhaps one or two lone wolves did. |
|
On February 9, 1959, after a fresh snowfall, the first census was made. A pack of 15 wolves was discovered near McCargo Cove, a lone wolf at Todd Harbor, and another individual in Rock Harbor. A search of the rest of the island produced no other wolf sign. However, on February 23 a pack of 3 was sighted near Five-Finger Point, and a few minutes later the pack of 15 plus a lone wolf were discovered near Davidson Island, demonstrating that at least 19 wolves were present. The extra lone wolf seen February 9 might have been a straying member of the pack of three. No other wolf sign was seen in 1959 which could definitely be attributed to any other animals. In 1960, four groupings of wolves were noticed: 15 (plus a lone wolf which followed this pack closely), 3, 2, and 1. The two wolves were seen three times, and the pack of three, five times, all on the same half of the island, but both packs never were observed on the same day. Therefore, I thought that perhaps the two wolves were part of the pack of three, and that the single wolf (only noticed once that year) was the third animal. The total estimate remained at 19 or 20. |
 Figure 34—Tracks of five wolves in sand. |
 Figure 35—Wolf tracks in snow. Therefore, the 1961 estimate of the number of wolves present on Isle Royale is 21 and possibly 22. The difference between estimates in 1960 and 1961 is caused only by the difference in interpretation of the observations. That the pack of two was not seen in 1959 does not mean it was not present, for during that year even the pack of three was observed only once, whereas in subsequent years it was seen many times. I believe that the Isle Royale wolf population has remained unchanged for the duration of this study. My increasing familiarity with the island's wolves from one study period to the next merely has made the last census most precise. |
The census in 1961 was complicated by the fact that the large pack often split up. Nevertheless, this pack still contained 15 animals. Lone wolves and the pack of three again were sighted several times. This year, however, strong circumstantial evidence indicated that an additional pack was present, composed of two animals. Although this never was proved conclusively, general knowledge of the Isle Royale wolves makes me believe that the group of two animals seen in 1960 and 1961 was not part of the pack of three.
|


Packs
A summary of the size of Isle Royale wolf packs observed before and during this study is presented in table 6. Because larger packs sometimes break up into smaller groups, single observations are not always reliable for determining the size of a pack. Nevertheless, it is interesting that sightings recorded before 1959 involved some groupings of the same size as those seen during this study.
TABLE 6.—SIZES OF WOLF PACKS OBSERVED ON ISLE ROYALE
| Year | Sizes of packs | Source | |||||
| 1 | 2 | 3 | 4 | 7 | 15-16 | ||
| 1952 | .. | X | .. | .. | .. | .. | Hakala (1954)a |
| 1953 | X | .. | .. | X | .. | .. | Cole (1953)a |
| 1956 | X | X | .. | X | X | .. | Cole (1956)a |
| 1957 | X | .. | X | X | X | .. | Cole (1957)a |
| 1959 | X | .. | X | .. | .. | X | Present studyb |
| 1960 | X | X | X | .. | .. | X | Present studyb |
| 1961 | X | X | X | .. | .. | X | Present studyb |
bThese figures represent basic pack sizes; temporary groupings are not included.
Hakala (1954) observed two wolves along the Feldtmann Trail in 1952, and Cole (1956) found evidence of a pair in 1956 near Siskiwit Bay. In 1960 and 1961, a pair (one animal larger than the other) was observed only on the northwest side of the island. Perhaps a pack's territory changes as variations occur in the size or distribution of other packs in a discrete population. If that is so, this pair may be the same as that observed in previous years.
Each winter a pack of three frequented the northeast and northwest parts of the island, where Cole (1957) four times observed a group of three. Probably these are the animals observed by Cole. One of the members is smaller than the others, so it may be a female.
The most significant pack on Isle Royale usually contains 15 to 16 members. This pack probably represents some combination of the seven wolves and the four observed by Cole, and the offspring of either or both groups. It has the largest (and probably best) range, kills the most moose, and dominates in encounters with other wolves. Most of each winter study period was devoted to observing this pack.

Figure 38—A typical formation of the large pack.
During the winters of 1959 and 1960, members of the large pack usually remained closely associated (figure 38). The few times the pack did split up in 1959, it usually separated into groups of 10 and 5 to 6. The smaller group sometimes continued to rest for about an hour after the other animals started traveling, and it occasionally lagged on long treks. Twice, 5 wolves headed for old kills while the other 10 continued hunting. (In August 1960 and May 1961, tracks of a pack of five were observed along the winter runway usually used by the large pack.) Within the group of five, three animals seemed lighter colored and lankier, and these were thought to be young-of-the-year. If they were, this might explain why the group rested longer and more frequently.
 Figure 39—Moose trail used often by wolves in summer. |
In 1960 the large pack still contained 15 to 16 members, but the 3 lanky wolves were not evident among them. The only time we observed any break-up of the pack was during the last 4 days of the study period, March 17 to 20, when three animals were missing.
However, in 1961 this pack was split about half of the time. On 13 of the 25 days the pack was observed, it was divided, usually into 5 and 10, or 7 and 8. There also were indications that it might have been losing a member, for often when the groups were united, only 14 wolves were present. In Alaska, Burkholder (1959) studied a pack which usually numbered 9 or 10 but sometimes split into 3 and 7.
Within the large pack there appeared to be at least three females, as determined by their behavior during the mating season. One of these, the smallest individual in the pack, was accompanied closely by a large male for a few weeks each winter. This pair, part of the 10 when the pack split in 1959 and in 1961, was the only pair that was consistently evident in the pack.
At least one lone wolf has been seen each year of the present study and of Cole's studies. In 1957 one followed Cole and his pilot for 9 miles across Siskiwit Bay. Cole (1957) believed this may have been Big Jim, the tame wolf released in 1952.

Figure 40—View toward Canada
(Sibley Peninsula, Ontario, 20 miles in background) from interior of
Isle Royale.
The lone wolf studied during the present investigation followed the pack of 15 from February 23 at least to March 14, 1959. Usually, it remained about 100 yards behind the pack and often was chased. Throughout the 1960 winter study period, a lone wolf (assumed to be the same one) again followed the pack, but it seemed almost to be accepted. It still traveled behind the others and did not mingle much, but only on February 22, when much mating activity occurred, did I see it run from them. On that occasion, whenever they looked or moved toward the animal, it ran and then followed the group from a distance. The relationship between the pack and this individual in 1960 is difficult to describe, but it seemed more a matter of strong tolerance by the pack than complete acceptance. Therefore, the basic size of this pack is considered 15 animals, although sometimes the "pack of 15 to 16" or the "15 to 16 wolves" will be referred to.
In 1961, two lone animals frequented the large pack's territory, and they could not be distinguished. Neither followed the pack consistently. One was probably the same individual seen in previous years, and the other presumably was the straying 15th member of the pack. Neither was exceptionally small, so both probably were males.
Other single wolves were seen each winter, but these may have been just straying members of the packs.
Home Ranges and Territoriality
Evidence that wolves are territorial was presented by Murie (1944) and Cowan (1947), and Stenlund (1955:37) wrote:
Travel routes [in Minnesota] suggested established home ranges with poorly defined borders overlapped somewhat by ranges of other packs. Home ranges from which the main pack had been removed remained barren of sign for two or three weeks after which other wolves reoccupied the range. Since these vacated home ranges remained free of wolves for a period of time, it is probable that wolves and wolf packs on the periphery respected the established territory of the home pack.
Schenkel (1948) believes that urine sprayed on scent posts serves to mark territories. This may explain Young and Goldman's (1944) report that urine from a strange wolf causes great excitement in other wolves and that scratching and kicking up of dirt, and often excessive deposits of excreta, are noted when such urine is found on scent posts.
Isle Royale's packs also seem to be territorial, at least in winter. Although the large pack used all parts of the park, it frequented certain sections much less than others. During 5 weeks in 1959, this pack only once visited the northwest shore from Duncan Bay to Thompsonite Beach, and spent but 4 days there. For the remainder of the period, the animals used the southeast side of the island (figure 41). In 7 weeks of 1960 the pack spent a few days on part of the northwest side, but continued to use the southeast half extensively (figure 42). However, in 1961 the range of the large pack seemed to have shifted somewhat. The southwest end of the northwest shore was used more, and the northeast end of the southeast shore, less. Even so, most activity of this pack occurred on the southwest end of the southeast side, as it had in 1959 and 1960 (figure 43). The shifts in range during the three study periods may have been apparent only, because throughout most of the year the wolves may have used many other regions than indicated in the figures.
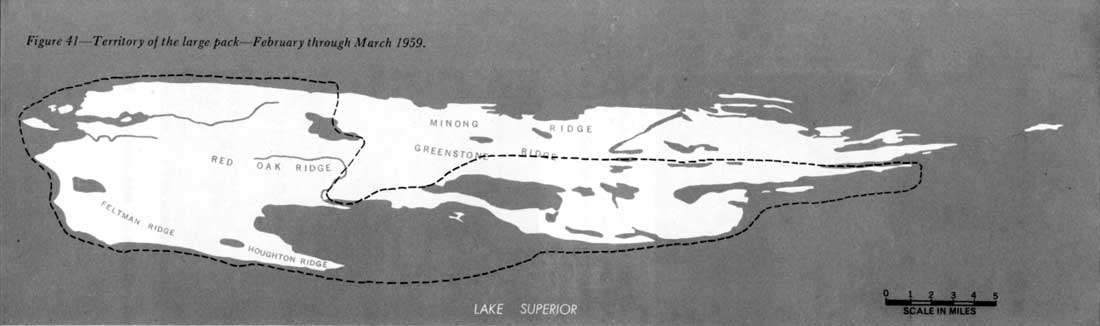
Figure 41—Territory of the large pack—February through March 1959.
(click on image for an enlargement in a new window)
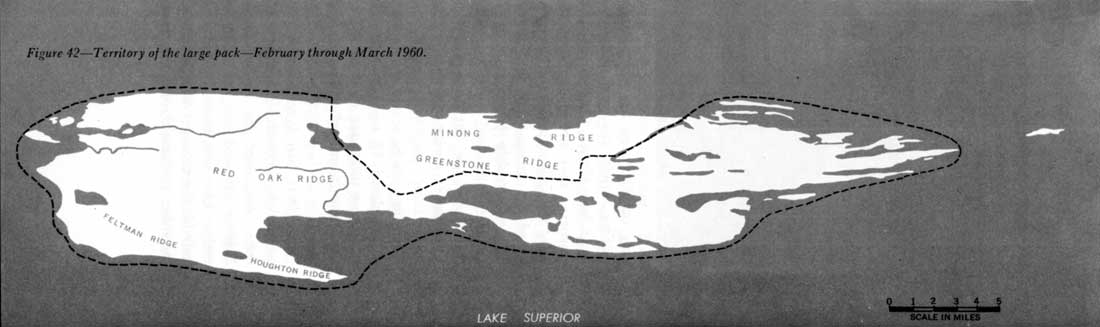
Figure 42—Territory of the large pack—February through March 1960.
(click on image for an enlargement in a new window)
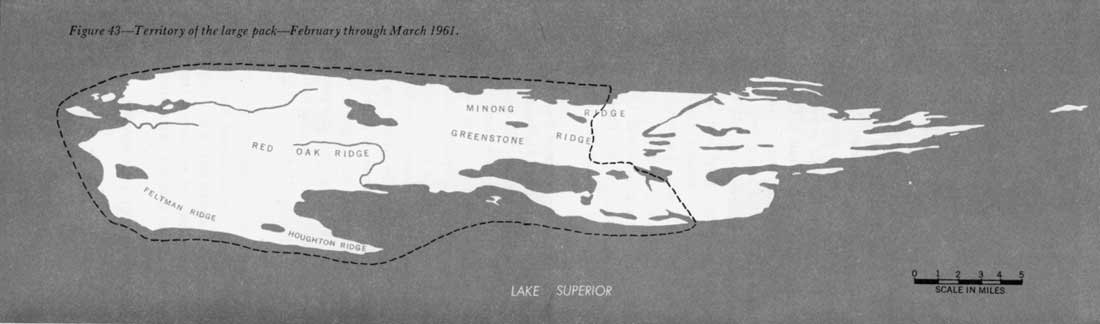
Figure 43—Territory of the large pack—February through March 1961.
(click on image for an enlargement in a new window)
The only summer observation of wolves which probably were members of this pack was reported by K. Knoble of Gays Mills, Wis. On July 17, 1958, he saw six wolves on the Huginnin Cove Trail. Tracks of a pack of five were seen in August 1960 and May 1961 along the southeast and southwest shore of Isle Royale, and a large group was heard howling north of Siskiwit Lake in June 1960. All these locations are within the winter range of the pack of 5. A large pack also was heard several times near Daisy Farm in Rock Harbor, a less-used section of the winter range.
Thus the winter range of the large pack could be considered to be about half of Isle Royale, or 105 square miles. The approximate density of wolves in this territory, then, would be one animal per 6.5 square miles. It probably is significant that this area contains the best moose range and about two-thirds of the winter moose population. Of course, it might be more appropriate to consider the whole island as the range of this pack, since the entire area is available to the wolves, and indeed they do occasionally visit all of it.
The pack of three and the pack of two frequent the northwest side of Isle Royale, apparently with complete coincidence of territories (figure 44). Four summer observations of three wolves were all made in regions within the winter territory of the pack of three. Campers saw three wolves at the head of Tobin Harbor about August 20, 1958, and Milford Johnsson observed three in October 1958, near Amygdaloid Channel. On July 28, 1960, Park Service Naturalists Robert A. Janke and Robert G. Johnson spotted three wolves just northwest of Mount Ojibway, and on August 20, 1960, Prof. and Mrs. W. Warth of Oberlin, Ohio, frightened three animals from a kill along the Huginnin Cove Trail.
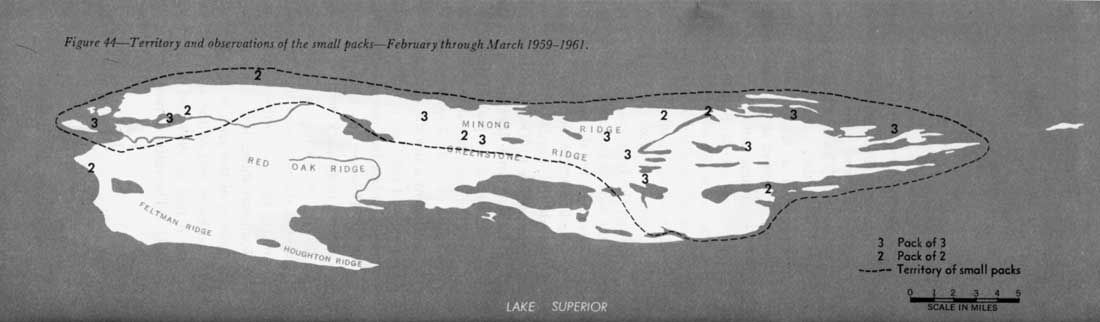
Figure 44—Territory and observations of the small packs—February through March 1959-61.
(click on image for an enlargement in a new window)
Since the two smaller packs range over about half of Isle Royale, the approximate density of wolves in their territory is one animal per 21 square miles, about one-third the density on the other half of the island.
The average approximate density for the entire island is one wolf per 10 square miles, but this figure should be viewed cautiously because of the disparity in densities between the two sections of the island. Wide differences exist among figures reported from other areas, but Isle Royale has one of the highest densities recorded. Stenlund (1955) estimated that northern Minnesota supports one wolf per 17 square miles. For three study periods in Saskatchewan, Banfield (1951) estimated densities of 39.5 square miles, 58 square miles, and 83 square miles per wolf. Cowan (1947) believes that in Jasper National Park there was one wolf per 87 to 111 square miles in summer, but one per 10 square miles in winter. Reported home-range sizes of individual packs are listed in table 7.
TABLE 7.—REPORTED HOME-RANGE SIZES OF INDIVIDUAL WOLF PACKS
| Location | Size of pack |
Sq. mi. of home range |
Sq. mi. per wolf |
Source |
| Wisconsin | 3 or 4 | 150 | 40 or 50 | Thompson (1952) |
| Michigan | 4 | 260 | 65 | Stebler (1944) |
| Alberta | 8 | 540 | 68 | Rowan (1950) |
| Northwest Territories | 7 | 90 | 13 | Banfield (1954) |
| British Columbia | 4 or 5 | 50 | 10 or 12 | Cowan (1947) |
| Alaska | 10 | 500 | 50 | Burkholder (1959) |
| Minnesota | 7 | 126 | 18 | Stenlund (1955) |
| Minnesota | 2 | 36 | 18 | Stenlund (1955) |
| Minnesota | 5 | 50 | 10 | Stenlund (1955) |
| Minnesota | 3 or 4 | 85 | 21 or 28 | Stenlund (1955) |
| Isle Royale | 2+3 | 105 | 21 | Present study |
| Isle Royale | 15—16 | a 105 | b 6.5 | Present study |
| Isle Royale (total) | 21 | 210 | 10 | Present study |
a If the entire island were considered to be the range, this figure would be 210.
b If the entire island were considered to be the range, this figure would be 13.
The winter range of the smaller Isle Royale packs overlaps with that of the large pack in the Rock Harbor and Washington Harbor areas. It is interesting to speculate whether the smaller packs chose to inhabit the portion of the island with proportionately fewer moose or whether they were forced there by the large pack. Murie (1944:44) wrote that ". . . it is advantageous for minor packs to find territories where they are unmolested." This may be especially important on Isle Royale because of the great numerical difference between the large pack and the smaller ones. Schenkel (1948:90), during a study of wolf behavior, concluded that ". . . as soon as the society controls a certain number of individuals, the manifestation of all individuals toward individuals from outside becomes more secure. . . ." Thus, it appears that the smaller packs probably have been forced to inhabit the part of the island in which they are least molested.
Two instances were observed of direct encounters by the large pack with other wolves. On February 7, 1960, the 16 wolves chased a single animal at least halfway across Moskey Basin (about one-half a mile) to the north shore of the bay. All ran extremely fast, but the pursued wolf outran the others. Upon reaching shore, it continued at top speed into the woods and then northeastward at least a quarter of a mile without stopping. The pack gave up when it reached shore, and the animals lay down and rested.
The second instance was observed on March 4, 1961, near Cumberland Point. The large pack was traveling along the shore from Rainbow Cove to Cumberland Point when two wolves, which had been feeding on an old kill, ran out of the woods about 125 yards ahead. The pack gave chase, and the larger of the two wolves headed into the woods and was not pursued. The smaller individual continued with utmost haste for a few hundred yards along the shore, stopped momentarily, looked back at the oncoming animals, and assumed the attitude of complete submission described by Schenkel (1948), i.e., front legs stretched forward and head and shoulders lowered. After a few seconds, it turned and headed along the shore, with the pack in continued pursuit. All ran swiftly, but the pack did not gain on the lone wolf. The pursuers stopped after covering about one-half a mile; the single wolf continued at the same speed for at least another mile before entering the woods.
There also were indirect indications of enmity by the large pack toward outside wolves. On March 6, 1960, the pack of three ran "anxiously" from Grace Island to Washington Island while the 16 wolves were heading across Grace Harbor, a quarter of a mile away. Grace Island prevented each pack from seeing the other, but the 16 animals kept looking toward the 3, which were running and watching their back-trail. The large pack did not pursue. On February 22, 1960, the large pack was traveling overland south of Ishpeming Point when suddenly half of the wolves struck out on a fresh wolf track. They followed it excitedly for about a quarter of a mile before returning to the others.

Figure 45—Part of large pack traveling along
shore of Washington Harbor.
The actions of the pack of 15 to 16 toward other wolves gives the impression that if the dominant animals ever caught the outsiders, a mortal fight would ensue. Indeed, Cowan (194?) reported an instance related to him in which a large wolf was found mangled by others, and another instance in which four wolves attacked a fifth and wounded it badly. He also wrote of a situation in which a wolf wounded a dog and then rushed it again as its master leaned over it. Cowan believes the wolf's action was a manifestation of territoriality. Murie (1944:43) also described an observation of a wolf pack wounding an alien wolf. On Isle Royale, Cole (1956) found tracks indicating that a pack had attacked a strange wolf. Bloody snow and a 2-inch piece of lip showed that a serious fight had ensued.
Movements
"The desire to travel appears to be an inherent trait in wolves" (Stenlund, 1955:30). This statement Seems to apply well to wolves on Isle Royale, for they often travel long distances, bypassing areas with high moose concentrations, and sometimes doubling back on their own tracks before making a kill. Kelsall (1957) analyzed 71 wolf observations involving 2,552 minutes, and found that 34 percent of the wolves' time was spent in traveling. Although no such figure was sought during the present study, indications are that Isle Royale wolves probably spend a comparable amount of time traveling.
Much travel seems to be necessary to the island's wolves for locating susceptible prey. Once they consume a carcass, any moose they detect is subject to attack, but before encountering a vulnerable animal, the wolves may travel 60 miles or more (table 8). Burkholder (1959) in Alaska found that distances between kills made by the pack he studied varied from 6 to more than 45 miles, and averaged 24.
TABLE 8.—DISTANCES (MILES) TRAVELED BY LARGE PACK BETWEEN KILLS
| Year | Minimum | Maximum | Average | Number of observations |
| 1959 | 0 | 60 | 30 | 9 |
| 1960 | 10 | 67 | 27 | 11 |
| 1961 | 6 | 44 | 19 | 5 |
| 3 years | O | 67 | 26.5 | 25 |
It is well established that wolves travel where going is easiest. In winter they follow frozen rivers, lakes, and streams; open ridges; and hard-packed drifts. Isle Royale wolves use such features also, but they follow the shoreline most extensively. There the snow is wind-packed, and the footing is good. Travel habits similar to those of the island's wolves were noted by de Vos (1950:174) in wolves on nearby Sibley Peninsula, Ontario:
. . . in late winter and early spring wolves travel extensively on the ice along the shores of lakes. They may either follow the shoreline into bays or cross those in a straight line. Often they run from land point to land point or from one small island to another in the bays around the peninsula.
He concluded that travel routes are determined by topography, distribution of prey, and seasonal changes. Stenlund (1955) stressed the importance of topography, and this factor seems most significant on Isle Royale also.
Isle Royale wolves usually travel single file in winter, especially during overland forays. This appears to be a common habit of wolves, for it has been reported often. Not only is this mode of travel more efficient, but the packed trails that result become convenient overland travelways for the future. Regular use of such a runway keeps it easy to travel despite a heavy accumulation of snow.
Although the wolves commonly use the same trails whenever they pass through an area, they do not have a predictable travel routine. This agrees with work by de Vos (1950) and Stenlund (1955). The island wolves usually do not even follow a circuitous route, although circuits of runways do exist. Most authors agree that wolves follow their circuits in both directions. The Isle Royale animals are no exception, for they often double back on their own tracks.
Established wolf trails are used year after year on Isle Royale, just as they are in other wolf ranges. Many of the trails reported by Cole (1957) were still used during the present study, but less-used side trails varied from year to year. Side trails seem to originate as routes used by wolves in pursuit of moose. Once established, they may be used several times in winter. However, we observed a few occasions when wolves struck out overland without resorting to old trails. Stenlund (1955) also found this in Minnesota. In British Columbia, Stanwell-Fletcher (1942) tracked a pair of wolves which plowed chest-deep for 22 miles in 6 feet of fluffy snow, without lying down to rest.
We were notable to follow either the pack of two or the pack of three for more than part of a day, so extensive information on their movements was not obtained. The north shore from Washington Harbor to McCargo Cove probably was the route used most, but a trail was sometimes found from McCargo Cove to Blake's Point and around into Rock Harbor and Moskey Basin. An alternate route from McCargo Cove to Moskey Basin followed the chain of lakes from Chickenbone Lake to Lake Richie. The Minong Ridge from McCargo Cove to Todd Harbor was used often, and the Greenstone Ridge Trail, packed by moose tracks, some times was followed (figure 39).
In summer, wolf tracks and scats have been found frequently on all Park Service trails within the winter range of these packs, so I assume that these trails constitute major summer routes (figure 3). The Minong Ridge and the extensive system of moose trails also are used in summer.
The most-used winter route of the large pack followed the south shore from Washington Harbor to Halloran Lake and Siskiwit Bay, or to Houghton Point, then across Siskiwit Bay (or around its periphery), and along the shore to Malone Bay. From there one route cut across to Siskiwit Lake, Intermediate Lake, Lake Richie, and Rock Harbor; another followed the shore to Chippewa Harbor and then crossed to Rock Harbor (figure 47).
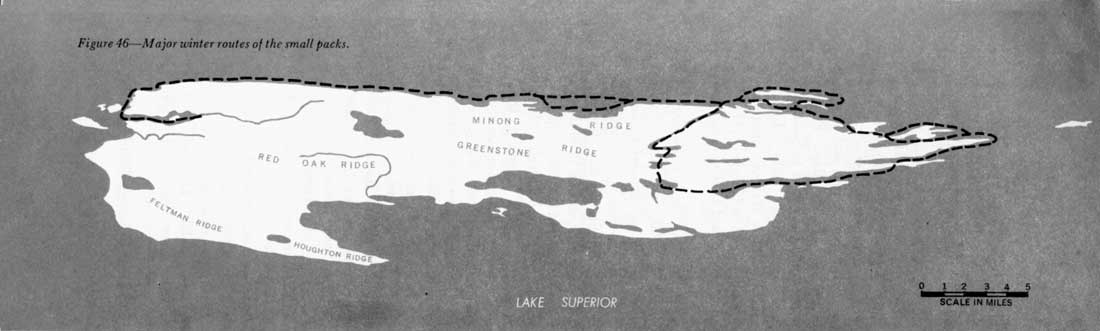
Figure 46—Major winter routes of the small packs.
(click on image for an enlargement in a new window)
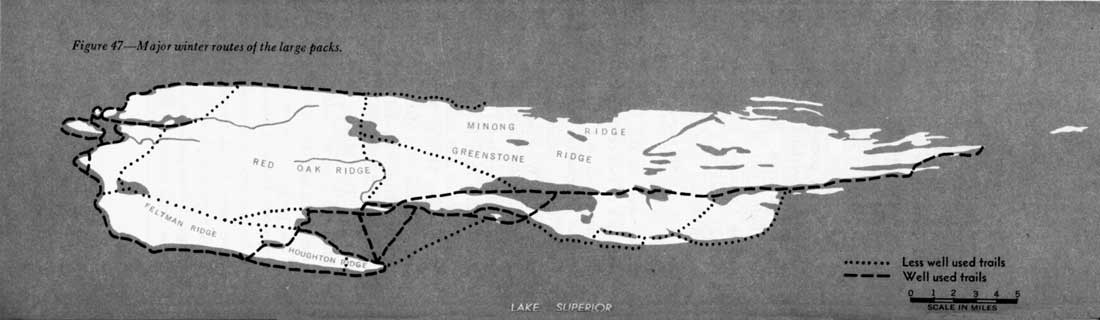
Figure 47—Major winter routes of the large packs.
(click on image for an enlargement in a new window)

Figure 48—Two members of the large pack.

Figure 49—Ten of the large pack file through deep snow.

Figure 50—One member of the large pack runs when author steps out
of nearby shack.
During 31 days, from February 4 to March 7, 1960, when the entire route of the 16 wolves was known, the animals traveled approximately 277 miles, or 9 miles per day (figures 51—55). However, during 22 of those days the wolves fed on kills, and no extensive movement occurred. Thus, in 9 days of actual traveling, the animals averaged 31 miles per day. During the entire study, the longest distance known to have been traveled in 24 hours was approximately 45 miles. In Alaska, Burkholder (1959) followed a pack that traveled a maximum of 45 miles in a day and averaged 15 miles per day for 15 days' travel, presumably including feeding periods. In Minnesota a pack moved 35 miles overnight (Stenlund, 1955).
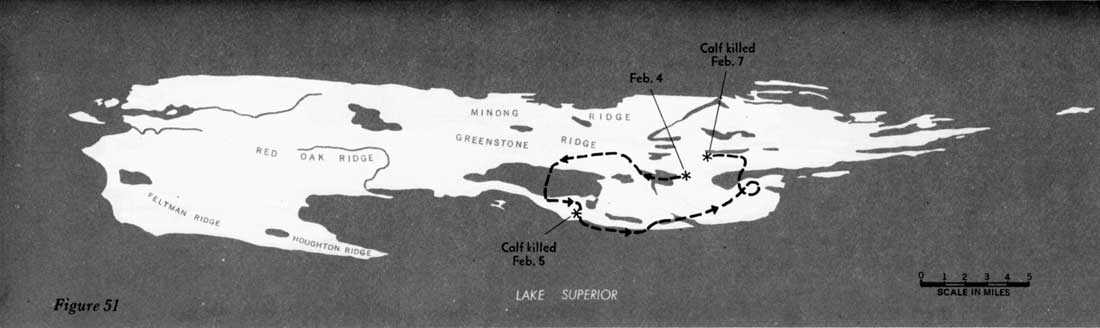
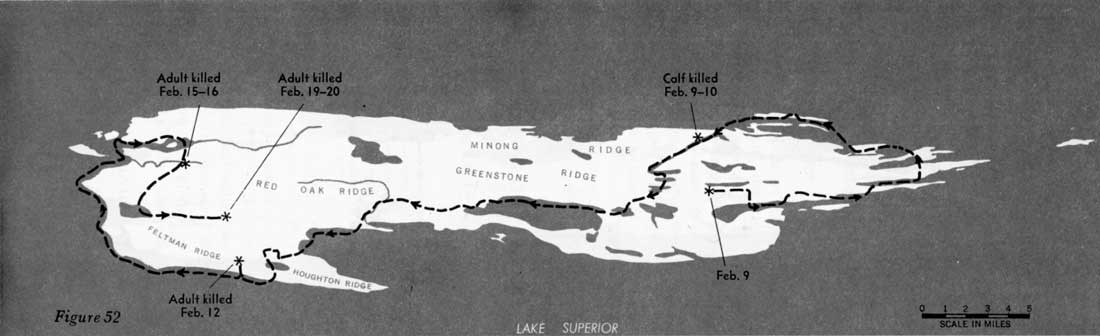
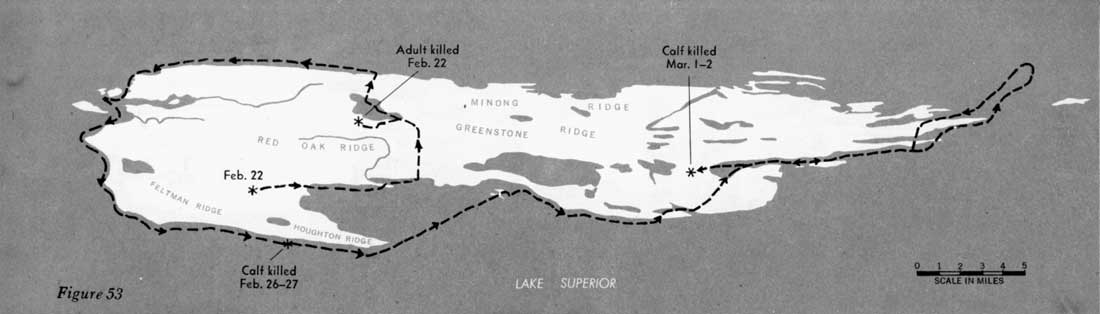
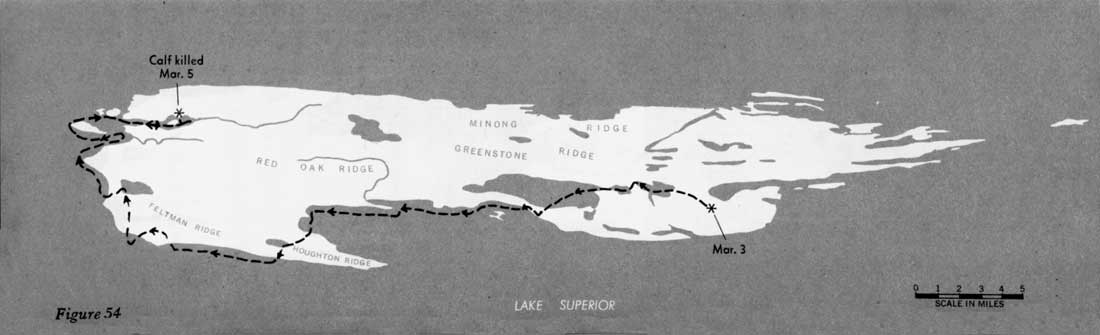
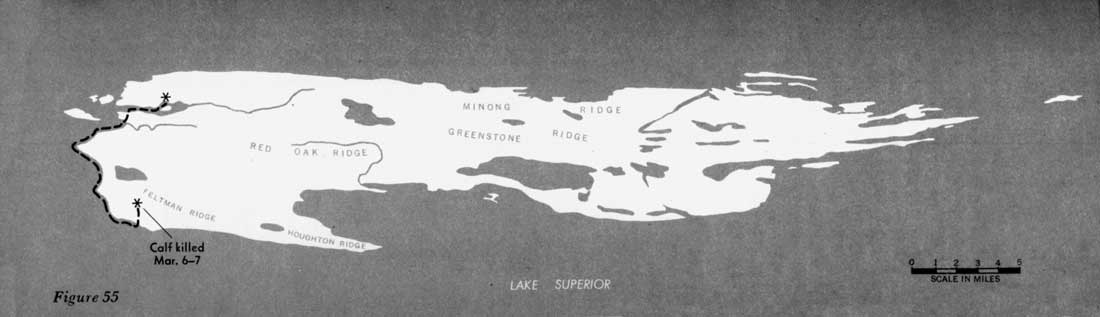
Figures 51—55—Routes of large pack
February 4 through March 7, 1960.
(click on images for an enlargement in a new window)
The wolves usually travel at a trot, about 5 miles per hour. They rest every few miles, especially on the day after leaving a kill. Generally they leave soon after dawn and begin in summer the large pack continues to use many of its winter routes.
Most lone wolves were seen along routes used frequently by the packs. but, of course, some of these animals may have been strays from the packs. Banfield (1951) also found single wolves following routes used by packs.
Social Behavior
Although few opportunities existed for studying at close range the actions of the Isle Royale wolves, certain behavior was noticeable from the aircraft. No attempt was made to use foreign urine, dummies, or howling records for analyzing the actions of the wolves, but these techniques are suggested for future workers. The behavioral information presented is based upon distant observation of undisturbed wolves.
ORGANIZATION OF THE PACK
According to Young and Goldman (1944:120):
The pack is generally a pair of wolves and their yearling or two-year-old off spring. At times, however, there will be an intermingling of several wolf families to form a pack; but the duration of such bands is short.
Olson (1938) also asserted that packs are family groups and that larger packs consist of two or more families. Murie (1944), through recognition of individual wolves, as certained that the "family theory" of the pack held for two groups of wolves in Alaska. He observed members of the East Fork family together at various times from May 15, 1940, to March 17, 1941. In 1941 two females from this pack had young, each in a separate den, but by June 30, one group had moved in with the other. Another pack, seen in August and in December, each time contained the same wolves, three adults and six pups.

Figure 56—Part of large pack traveling across ice.
Theories conflict regarding the status of the pack in summer. Schenkel (1948) believes that each mated pair leaves the pack toward winter's end, the unpaired, weaker, and younger animals staying in small groups for some time. Cowan (1947) also thinks that in summer the hunting packs are broken up and that the animals hunt in smaller groups. However, Murie observed 15 wolves (including 2 litters of pups) in one group in July. He also saw five adults at a den in June and July. Whether members of a pack dissociate in summer may depend upon the type of terrain, the prey species, the composition of the pack, and several other variables.
On Isle Royale, most spring, summer, and autumn observations have been of single animals, which may have strayed, at least temporarily, from packs (table 9). Tracks, howling, and sightings of three associated wolves in summer and autumn indicate that probably the pack of three functions as an entity most of the year. A larger group also was heard howling several times. This, plus an observation of a group of six adults, and tracks of five adults show that on Isle Royale some wolves associate with others during the summer, at least at times. It may be that bonds among adult wolves are stronger in winter but that members of packs are together frequently in summer.
TABLE 9.—NUMBER OF WOLVES SEEN OR HEARD FROM MAY TO OCTOBER
| Numbera | Date | Location | Observer |
| 6 | July 1958 | Huginnin Cove Trail | Reported |
| 3 | Aug. 1958 | Head of Tobin Harbor | Reported |
| 1 | Aug. 1958 | Rainbow Cove | Reported |
| 3 | Oct. 1958 | S. shore of Amygdaloid Channel | Reported |
| 1 | May 1959 | Lake Desor | Author |
| 1 | May 1959 | Hay Bay | Reported |
| 1 | June 1959 | Sugar Mountain | Author |
| 1 | July 1959 | Daisy Farm | Reported |
| 1 | July 1959 | Lake Desor | Author |
| 3 | Oct. 1959 | Mt. Ojibway | Author (heard) |
| Several | May 1960 | Daisy Farm | Author (heard) |
| Several | May 1960 | Daisy Farm | Author (heard) |
| 3 | May 1960 | Daisy Farm | Reported (heard) |
| 1 | May 1960 | Mt. Ojibway | Reported |
| Several | June 1960 | N. of Siskiwit Lake | Author (heard) |
| Several | July 1960 | Daisy Farm | Author (heard) |
| Several | July 1960 | Daisy Farm | Author (heard) |
| 3 | July 1960 | Mt. Ojibway | Reported |
| 2 or 3 | July 1960 | Conglomerate Bay | Reported (heard) |
| 5 | Aug. 1960 | Attwood Beach | Author (tracks) |
| 3 | Aug. 1960 | Huginnin Cove Trail | Reported |
| 3 or 4 | Aug. 1960 | Chicken bone Lake | Author (from air) |
| 4 | Aug. 1960 | Chickenbone Lake | Author (heard) |
| Several | Aug. 1960 | Daisy Farm | Author (heard) |
| 5 | May 1961 | Long Point | Author (tracks) |
| 2 or more | May 1961 | Malone Bay | Author (tracks) |
| 3 | May 1961 | Conglomerate Bay | Author (tracks) |
a In any of these cases, more animals could have been nearby but unobserved.
Murie (1944:45) explained what might cause a large pack to break up:
The size of the pack may be limited by the law of diminishing returns. Beyond a certain size, advantages may disappear. A pack might be so large that, after the strongest members had finished feeding on a kill, there would be little or nothing left for the rest. In such a situation, hungry ones would go off to hunt again, and the strong ones, already fed, would remain where they were. There thus might result a natural division of a band which was too large to function advantageously for all its members. One would expect that where game is scarce the wolves would operate in smaller units than where food is abundant.
The reason wolves form larger packs in winter seems to be unknown. Possibly, much more food is required during this season, and a larger pack might hunt more efficiently. The latter assumption might not be valid on Isle Royale, for while the large pack chases a moose, usually only five or six animals stay close to it; the others fall far behind. Because of this, it seems that the most efficient pack would contain five or six animals. Two packs of this size, operating independently, could travel twice as far as one, and therefore could locate, on the average, twice the number of vulnerable moose. It may be significant that in 1961, when fewer calves were present, the large group was split into two packs about half the time. Since calves provide much of the winter wolf food (about half during the 1960 study period), a shortage of this age class could cause more difficult hunting, which might force the wolves to operate in smaller groups. Indeed, in the 1961 study period when the large pack split up several times, the 15 wolves consumed more food than in either of the previous two periods.
At times, a larger pack might be more advantageous. Many a moose stands its ground when cornered by wolves. Such an animal usually is safe, for if the wolves cannot force it to run, they soon leave. Possibly a moose is more inclined to flee when confronted with several wolves than with few. If that is true, a larger pack would be more advantageous, for when a moose runs, it is much more vulnerable.
In the Rocky Mountain national parks of Canada, "the usual winter hunting pack consists of from four to seven individuals, with five or six the most frequent numbers. Packs of 10 or 12 have been reported once or twice in the Jasper area. The largest group recorded was believed to contain 14 individuals" (Cowan, 1947:157). Stenlund (1955) reported that a pack of 15 occurred in Minnesota, and Olson (1938) gave records of packs containing 20 and 30 wolves, also in Minnesota. Murie mentioned a sighting of 22 wolves, tracks of 24, and a report of 50, in Alaska.
According to Schenkel (1948:83) packs are formed in early winter. He provides the following description:
Chorus, howling, joint wanderings and hunting, and fairly early rivalries concerning leadership and sexual partnership denote this period. During this time the pack becomes a close (exclusive) society. Its core comprises the bitch wolf, presumably the only mature one of the pack, and the male "lead wolf." Whether the isolation of the mature female wolves from one another is the result of rivalries, what course these rivalries take in any event, and what effect they have on the formation of a pack is not known. The lead wolf and bitch more and more plainly be come a pair—first in the pack group—then at winter's end they separate from the pack and occupy a family area for the summer.
The possible beginning of a break up in the large pack was noticed in mid-March 1960. The 16 wolves had remained together from February 4 to March 16, but on the 17th and 18th only 13 were seen. On these dates the animals were inland and could have been miscounted, but on March 20 (the last day of the study period) they crossed Siskiwit Bay; only 13 were present. Perhaps this disbanding was only temporary, but it might have been the beginning of a seasonal breakup.
SOCIAL RANKING WITHIN THE LARGE PACK
In the zoological gardens where Schenkel studied wolf behavior, more than one mature male and female were present in a pack, but one of each sex was dominant. These two highest ranking individuals he called "alpha animals." Murie (1944) believed that an unmated male was leader of one of the Mount McKinley packs, for other wolves approached this individual cowering. Apparently, this animal was dominant even to the mated pair within the pack.
Isle Royale's large pack contains at least three mature females, but only one pair (a male and female closely associated for 2 or 3 weeks) was observed each winter. On February 19 and 22, 1960, the female led the pack, while the male remained beside her, half a body length behind. Copulation was attempted several times. The female appeared inexperienced at leading, for she backtracked twice and was often shortcut by other wolves. Each time after shortcutting the lead pair, the rest of the pack waited "respectfully," and as the leaders passed, each individual assumed the submissive position described by Schenkel (1948:fig. 2). While passing these animals, the leaders held their tails high, in the dominant position (Schenkel, 1948:fig. 30a).
On February 6, 1961, the small female also led the pack, followed closely by a male. Both held their tails in the dominant position when approaching an old kill, whereas the rest of the animals held theirs normally. This pair probably was the "alpha" pair. Apparently, the male usually was leader, but while his mate was in estrus, he took the advantageous position behind her. Fuller and Novakowski (1955) reported that during a wolf-poisoning campaign in Wood Buffalo National Park, Canada, males dominated in taking the bait in two instances in autumn, whereas the only instance during the mating season showed that a female was dominant.
The alpha pair does not always head the string of wolves, but position in line does not necessarily reflect social order. In Alaska, Murie (1944) noticed that the first animal in line was not always the leader. In the present study there were three main activities during which one wolf appeared outstanding: journeying overland, hunting, and arousing the pack from its rest. (I do not know if this wolf is the alpha male as identified above, or even whether it is the same individual each time.) When the pack journeyed through deep snow, the first wolf in line frequently was 25 to 50 yards ahead of the rest, even though it broke trail. When this animal rested, the others did also, and when it began to travel, the others followed.

Figure 57—Social behavior among members of
the large pack. Note oral "besnuffing" in uppermost pair, and varying
tail positions. The smaller wolves (arrows) maybe females.
During hunts in 1959 and 1961, one wolf often seemed more aggressive. Sometimes one animal threatened a moose while the others paid no attention. During several chases, one wolf caught up to the moose before the others did, and in some instances an individual continued chasing for 100 yards farther than the others. In a few cases, although several wolves threatened a moose, only one actually attacked the animal. Once when most of the wolves were lying around waiting for a wounded moose to weaken, one individual, its front legs covered with the blood of the prey, continued to harass the belligerent moose.
A hunt on February 12, 1960, produced some unusual behavior of a different type. After a three-quarter-mile chase during which most of the wolves ran alongside a moose for about 300 yards without attacking it, the lead wolf suddenly stopped, turned around and lunged at those behind, as if to stop them from continuing the chase. It succeeded, for the other wolves turned and ran up their backtrail.
One wolf usually arouses the rest from their slumber. After stretching, this animal goes from wolf to wolf, touching noses and awakening each individual. As each wolf arises, it duplicates the procedure until the entire pack is active. Perhaps it is not necessarily the leader which initiates the arousing; it could be the first animal which awakens. (Murie describes similar arousing behavior begun by an individual other than the leader.) Nevertheless, I frequently saw the leader begin such activity.
Without identifiable individuals in the pack, it was impossible to distinguish the dominance position of the middle-ranking animals. However, status-demonstration was observed often among the wolves in the large pack. Frequent urination, oral, anal, and genital "besnuffling" (figure 57), presentation and withdrawal of anal parts, tail wagging, and mock attack upon weaker members of the pack (an energy displacement) were evident almost every day the pack was observed. Schenkel (1948) describes the above behaviorisms and explains their significance. The large pack performed the most noticeable social behavior in the following situations: (1) upon awakening, (2) when stopping to rest, and (3) while reassembling after a hunt or after splitting up.
Schenkel (1948:87) explains why this behavior occurs so frequently:
Every mature wolf has an ever ready "expansion power," a tendency to widen, not necessarily his personal territory, but rather, his own social behaviour freedom, and to repress his "Kumpans" of the same sex. Consequently, he maintains a constant watchful interest in all socially important happenings within the pack. In particular, status quarrels are never private affairs between two individuals; the whole society takes a more or less active part in them.
Murie (1944), Young and Goldman (1944), and Crisler (1958) also have observed one or more of the social behaviorisms.
The most repressed individual in the large group of wolves was the most conspicuous. This was the lone wolf which followed the other 15. We first saw the animal on February 23, 1959, about 100 yards behind the pack. It held its head low, ears back, and tail between its legs in the submissive position, and appeared to fear the other wolves. On February 24, while the pack rested on a lake, this animal was able to join the last two wolves in line. These slept for long periods and were not interested in the mating activity in which the rest engaged. The lone wolf wandered around near these two, and suddenly two others bolted toward it. The low-ranking individual ran off, directly past the two friendlier animals, but these remained unconcerned.
After a 100-yard chase, the pursuing wolves cornered the lone wolf near a snowbank and attacked it. They fought the animal for a few seconds and then, rejoined the excited pack. The lone wolf followed slowly, and again the two wolves attacked it momentarily. This happened a third time, after which the lone wolf did not attempt to follow the attackers. The details of the fights were not observable, but the lone wolf fought hard and did not appear to be injured. Lorenz (1952) asserts, on the basis of observations in the zoological park, that a wolf submits to its aggressor by presenting its throat, a maneuver which tends to inhibit the aggressive tendency in the attacker. Perhaps this is what caused the hostile animals to end their attacks so suddenly.
The lone wolf continued to follow the pack for the remainder of the 1959 study period and throughout the 1960 winter study period. We did not notice any more attacks on the animal, but it stayed away from most of the wolves and remained submissive. Schenkel reports that "energy displacements" directed at subordinates often occur in packs, and may take the form of ambushes, sneak attacks, and fights. He often saw cases in which several wolves directed their attack against one animal over a long period. This individual "... steadily lost the significance of environmental social partnership, was robbed of all social initiative and, in certain circumstances, with repeated attacks, became mortally wounded" (1948:88).
Despite the hostilities shown it by certain members, the lone wolf seemed to be accepted by part of the pack. The two friendly wolves mentioned above provide one example. On another occasion, when most of the animals were resting, two wolves backtracked around a point about 25 yards to meet the lone wolf. They sniffed the cowering individual a few seconds and accepted it. The three then moved back around the point a few yards to the rest of the pack. When the lone wolf saw the pack, it ran about 25 yards, lay down, and remained there. A third case of differential behavior toward this individual occurred on March 4, 1959. Ten of the wolves started traveling, while the other five (including the three lighter-colored, lankier animals) rested. The lone wolf joined these five. When they left, it accompanied them and was completely indistinguishable from them; no trouble ensued. A similar situation occurred on March 8. Six wolves, including the lanky individuals, were several miles behind the rest of the pack. The lone wolf joined these and accompanied them, without any apparent fear, to the rest of the animals. When they approached the main pack, the usual sniffing, tail wagging, and other greeting behavior took place; but the lone wolf quickly ran off and remained away from the others.
The reasons for the differential reactions to the lowest-ranking individual are unknown. Possibly, each of the members which accepted it also held low social status. Indeed, there were reasons to conjecture that the three lanky individuals were pups.
Although the lone wolf was suppressed by the large pack, it joined the group in pursuit of a strange wolf. All 16 animals chased the alien for half a mile. According to Schenkel (1948), this type of behavior is not unusual. He observed that despite the sometimes-violent relationships within a pack, the members present a united front toward aliens and become a unit during friendly activity such as chorus bowling.
HOWLING
The full significance of wolf howling is unknown. Murie (1944) described situations in which several animals howled before leaving the den area to hunt. From his descriptions, it appears that the howling was merely a manifestation of the wolves' restlessness. This seemed to be the case also during an observation of Isle Royale wolves. On February 9, 1961, four wolves lay on the ice at the head of Washington Harbor from 8:45 to 9:15 a.m., after which they headed into the woods. At 10 a.m., a wolf howled twice from the woods near the shore, and a few minutes later an animal appeared on the ice and howled three more times. Each time, the wolf's muzzle pointed skyward; the howls were low-pitched and drawn out. Five minutes later, four more wolves appeared, one at a time. They walked about 150 yards onto the ice and lay down.
About 2 p.m. one animal arose, stretched, lay back down, and howled a few times, arousing the nearest wolf. Then it approached this individual, with tail straight up and tip cocked forward, and sniffed its nose. The second animal rolled over and extended its paws toward the first. Meanwhile, the other three wolves arose, and all five walked about 200 yards westward and disappeared up a creek bed. A few minutes later, a single wolf emerged from the woods, sniffed the tracks of the others, cowered, and howled for a few minutes. Then it wandered eastward along the shore for about 50 yards and entered the woods.
Crisler (1958:151) believes that howling is an emotional outlet for wolves. She writes:
Like a community sing, a howl is not mere noise, it is a happy social occasion. Wolves love a howl. When it is started, they instantly seek contact with one an other, troop together, fur to fur. Some wolves . . . love a sing more than others do and will run from any distance, panting and bright-eyed, to join in, uttering, as they near, fervent little wows, jaws wide, hardly able to wait to sing.
Seton (1937) and Young and Goldman (1944) believe that wolves vocalize when chasing prey. This supposition seems logical, for vocal expression might help keep members of the pack together as they chase their quarry. However, the only evidence I have found in the literature to support this contention was an observation in 1875 reported to Seton (p. 281) by a logger. During the present study, all hunting was observed from an aircraft, so only indirect information was obtained on the subject.

Figure 58—Pilot Don Murray and author examining a fresh
kill.
Two observations indicated that wolves did not vocalize while chasing moose. In one instance, the large pack chased and wounded a moose, while a second animal lay about 100 yards away. The latter moose eventually wandered away but did not seem cognizant of the 16 wolves nearby, as it probably would have if the wolves had been "tonguing." In the second situation, the large pack chased a moose, and a few of the animals caught up with it and held it at bay. Meanwhile, the others were wandering around searching for the moose. They finally found it by following the trail left by those which had cornered it. I believe that if any vocal communication had occurred, these animals would have run directly to the cornered moose.
One function of howling may be to aid in assembling. After long chases, the 15 wolves sometimes were scattered over a large area. On one such occasion, we noticed that a wolf ascended the nearest ridge and appeared to howl. Several others approached the first, and about 150 yards away another animal appeared to howl. Eventually, most of the pack assembled on the ridge. Murie (1944:102) described a similar episode.
On February 4, 1960, when we landed the aircraft within 100 yards of the 16 wolves on Intermediate Lake, the animals eventually scattered into the woods. A few minutes later we heard some howling, which soon increased in volume until the entire pack seemed to be involved. The whining, yelping, and howling (much of which was high-pitched) continued for about 30 seconds and then gradually diminished; a few single howls were emitted after the chorus had subsided. Tracks later showed that the wolves had assembled on a small knoll, where most of the howling probably originated.
Another time the pilot and I frightened the large pack from a freshly killed moose. As the wolves retreated, several barked hoarsely. We remained at the carcass for about 2-1/2 hours, and heard distant howling and barking intermittently throughout the period.
In the last instances some or all of the howling could have resulted from frustration or emotion. This undoubtedly was the case on an occasion in August 1960. From 9 to 11 p.m., I sat 20 feet up in a tree above a freshly killed moose. At 9:30 p.m. at least four wolves began howling about 200 yards away. Howling continued off and on for the next half an hour, but it gradually became more distant. A check the next morning showed that the wolves had not returned to the carcass.
Howling was heard several times near the Daisy Farm campsite (across Rock Harbor from my cabin) in the summer of 1960. The earliest time of day that I heard it was 5:40 p.m., and the latest, 12:30 a.m. It consisted of the usual medley of yips, barks, deep "mournful" howls, and extended calls of ever-changing pitch. Sometimes it occurred for only a few seconds, but once it lasted about 2 minutes. On one occasion when 22 campers were present at the camp site, several wolves howled directly behind the area. The animals sounded to me to be about 100 yards behind the lean-tos, although the campers thought they were closer. The reasons for all the howling in this area are unknown, but perhaps the sound of humans stimulated the wolves. Young and Goldman (1944) wrote that whistles and other human disturbances often stimulate wolves to howl.
Pimlott (1960) found that human "wolf" howling and recordings of wolf howling would cause wild wolves to perform. After extensive testing of this method he concluded (p. 7):
It appears that the stimulus of wolves to howl is, at least in part, directly proportional to the length of time since they last howled. It is frequently difficult to evoke a response within 15 to 20 minutes, or even longer, after they last howled.
Phonograph records of wolf howling were tried during the present study, and replies were obtained four times. The records were also played after "natural" howls, to determine whether the wolves would vocalize again within a few minutes of their first howl. Although only a few trials were made, results supported Pimlott's conclusion.
MISCELLANEOUS BEHAVIOR
Activity resembling play was noticed on March 6, 1960. The 16 wolves had just left a kill and were traveling along the shore toward Cumberland Point. Several animals chased one another back and forth and in circles, but sometimes a group would chase one individual and then suddenly turn on another. It appeared that the pursued animal carried something, possibly a bone, and that as it dropped the object, another would pick it up and attempt to out run the rest. The pack cut across Cumberland Point, and in the woods the activity reached its maximum. The entire pack became involved, some in ambushing, others in chasing, until eventually the animals tired. Several other times, I have noticed tracks indicating that the wolves had engaged in similar "sport."
Over-cautious behavior on the part of a single wolf was observed on February 28, 1961. At 6:05 p.m., seven wolves started northward across Hay Bay from Hay Point. Most of the ice was bare, and the wolves were reluctant to walk on it, probably because snow-free ice usually is new and thin. They tried to keep on the chunks of old, snow-covered ice, which were frozen together by new, bare ice. When there were no more snow-covered chunks, the wolves walked on the opaque cracks across the bare ice. However, one wolf would not follow the pack onto the snow-free ice, although it was tempted. Instead, it headed westward into Hay Bay on snow-covered chunks, being careful not to walk on bare ice. When the animal reached the end of these chunks and faced bare ice, it returned the 250 yards to where the pack had crossed.
Again the wolf started following the tracks, but once they left snow-covered ice, it would not continue. This time the animal ran about 150 yards southwestward, back into the bay where the snow-covered ice was continuous. It crossed this without hesitation. Meanwhile, the pack had reached shore and was about a mile away. The cautious wolf hurried to them, catching up at 6:35 p.m.
Reproduction
According to Schenkel (1948), pairing begins in early winter, and bonds strengthen as winter progresses. Winter rivalries within the pack occur only among members of the same sex, eventually resulting in an established social order. "In general, the usual conflicts of opinion remain somewhere in the middle between the two possible extremes (status demonstration—battle)" (Schenkel, 1948:88). However, apparently at times intense battles occur, for Crisler (1958:251) reported an instance in which one female killed another during a fight over a male. Regardless of the form rivalry takes, by mating season pairs are well established.
Young and Goldman (1944) reported that males mature in 3 years and females in 2. These authors (p. 84) provide the following account of wolf reproduction:
Wolves do not breed until between 2 and 3 years of age. They couple much as dogs do but can more readily separate. In captivity oestrum has been noted to continue from three to five days; the female has stood for the male over a period of five days, and then rejected further advances; not until the vulva became noticeably swollen would the female stand. The period of discharge of blood from its start in late December until the swelling of the vulva and the final copulation for five females averaged 45 days.
This places the actual breeding season at mid-February. A captive wolf, which Murie (1944) raised as a pup, first came into heat in early March of her second year, and remained in that condition about 2 weeks. Murie also reported on an other captive female, which failed to come into estrus the first year but bred with a dog the second year.
"The first 2 weeks that this wolf was in heat she fought off the dog but mated each day during the third week (March 9 to March 15). The male continued to pursue her on the following 3 days but there was no further mating after the fifteenth" (Murie, 1944:17). Four pups were born to this female on May 15, which establishes the gestation period at 60 to 66 days. Pups born in the Philadelphia zoo had a gestation period of 9 weeks (Brown, 1936). According to Bailey (1926) wolves in North Dakota bore young in March, so they must have mated in January. Murie (1944) reported that Mount McKinley wolves probably breed in early March, since young are born in early May. Fuller and Novakowski (1955), by examining female reproductive tracts, found that estrus probably occurred between March 5 and 21 in northern Alberta. Cowan (1947) believes that British Columbia wolves mate in March and early April.
I first observed mating activity among the Isle Royale wolves on February 21, 1959. This was the first day that the alpha pair was noticed. The male tried unsuccessfully to mount the female several times. One successful copulation was observed but probably not between these two wolves. When the animals coupled, the entire pack (strung out 100 yards ahead) raced back to the pair. After a few seconds of milling around, the pack left the two lying rump to rump. As we flew near the coupled wolves, they stood and snapped at each other but then lay down again. After 15 minutes they parted and hurried to the rest of the pack.
For the next half hour there were several attempted copulations between members of at least three pairs, but in each instance the female thwarted the male by sitting, tail between her legs. Each time mounting occurred, the nearby wolves rushed to the pair, in an apparent free-for-all. Schenkel (1948:93) presents a detailed description of precopulatory behavior in the wolf. I did not observe such behavior, but at the time, I was neither aware that it might occur nor close enough to notice it.
On February 24, much mating activity was evident, but only one successful copulation was observed, the animals being coupled back-to-back for at least 6 minutes. The last copulation witnessed in 1959 occurred on February 27, and lasted at least 8 minutes.
In 1960, complete coitus was observed only once, the wolves remaining coupled for at least 5 minutes. The activity of the rest of the pack indicated the presence of at least two other females. Chasing, fighting, and sniffing were noted on February 7, 14, 19, and 20; and on the 22nd, unsuccessful attempts at copulation were seen in one pair. No observations were made on behavior from February 23 to 29, but after the 29th no sign of mating activity was seen.
The only breeding behavior observed in 1961 occurred on February 6. During that day, much chasing and fighting (most evident during the mating season) took place. Flying conditions that year did not allow as much observation as in previous years. Nevertheless, the pack was observed for several hours a day on many days. Probably fear of the aircraft in 1961 caused the wolves to confine their breeding activity to periods when they were undisturbed (see p. 36).

Figure 59—Local snowstorms made flying treacherous.
No additional reproductive information was obtained on the Isle Royale wolves. Whether pups were born and raised is unknown. According to data presented by Stenlund (1955) and Fuller and Novakowski (1955), weight and size are not valid criteria for distinguishing adults from pups in winter. However, since sizes of the Isle Royale wolf packs have remained exactly the same for three winters, I believe that no pups have been added; it would be quite coincidental if exactly the same number of wolves died each year as were raised. As has been discussed, during the first winter, three lighter-colored, lanky individuals were observed in the pack of 15; these rested and played more frequently than the others and possibly were pups. They were not distinguishable in 1960 or 1961.
No active wolf dens were found, although much time was devoted to den hunting. However, on May 21, 1959, a freshly dug den was discovered on an open, south-facing slope about 350 yards north of Siskiwit Lake, opposite the western tip of Ryan Island. No fresh wolf sign was present, but the size of the entrance and tunnels indicated that probably wolves had dug the den. The entrance measured 28 by 17 inches, and the tunnels were 12 inches in diameter. The mound was 5 feet wide by 10 feet long. These measurements correspond well to those of wolf dens studied by Murie (1944), Cowan (1947), and Banfield (1954). In 1960, this den was partly caved in, but in 1961 it sheltered six fox pups (figure 60).

Figure 60—Fox pups at a den that once may have been a wolf den.
In 1960, Pimlott's (1960) method of locating wolf dens was tried. Recordings of wolf howling were amplified from 34 locations at various times between 7:55 and 10:15 p.m. (May 17 to August 5), and four replies were obtained. Three of these originated from the same location on the same evening (although broadcast sites were different), so only in two locations was contact established with wolves. Both areas were searched, but no sign was found. Since in each case no additional replies were obtained on the night following the first contact, it is probable that the replies came from traveling animals.
Food Habits
The timber wolf is a big-game predator. Smaller animals including birds, rodents, and lagomorphs are eaten, but I know of no wolf population which has thrived on small animals alone. Only one record was found which indicated that the majority of wolf scats from an area contained anything other than big game.
Tener (1954) reported that on Ellesmere Island 83 percent of 85 wolf scats contained arctic hare (Lepus arcticus) remains, whereas 17 percent contained muskoxen (Ovibos moschatus).
Murie (1944) found remains of caribou (Rangifer arcticus), Dall sheep (Ovis dalli), or moose in 935 of 1,174 wolf scats collected in Mount McKinley National Park; and big-game remains composed approximately 70 percent of the 1,350 food items. In eight wolf stomachs and eight scats from Michigan, deer and hare remains were represented equally (Stebler, 1944). Cowan (1947) reported that 80 percent of 420 wolf scats from the Rocky Mountain national parks of Canada contained remains of elk (Cervus canadensis), bighorn (Ovis canadensis), mountain goat (Oreamnos americanus), moose, caribou, or mule deer (Odocoileus hemionus). White-tailed deer remains occurred in 97 percent of 435 scats from Wisconsin (Thompson, 1952), and in 80 percent of 51 wolf stomachs collected in winter from Minnesota (Stenlund, 1955). Fuller and Novakowski (1955) found remains of bison (Bison bison) in 32 of 49 wolf stomachs from northern Alberta. Caribou remains composed 58 percent of the items in 62 scats from the Northwest Territories (Banfield, 1954).
On Isle Royale the moose population represents the only potential food supply which could support the present wolf population; beavers (except in winter) and snowshoe hares are available supplements. Of 87 scats collected in May 1952, 56 percent contained moose remains; 24 percent, snowshoe hare; and 20 percent, beaver (Cole, 1952a). In 1954, Cole reported that "sixty-five percent of the scats contained moose hair and 35 percent beaver hair," but the number of scats examined was not given.
A total of 438 wolf scats were analyzed during the present study (table 10). These were collected from 100 miles of foot trails in spring and summer from 1958 to 1960. Since at the time, it was not known whether coyotes were still present, only scats over 1 inch in diameter were considered wolf scats, in accordance with information presented by Thompson (1952). Whenever possible, scats were aged to the nearest month, and those of unknown age were designated "old."
TABLE 10.—ANALYSIS OF FOOD REMAINS IN 438 WOLF SCATS COLLECTED FROM TRAILS
| 1958 | 1959 | 1960 | Total | |||||||||
| Fresha | Oldb | Total | Freshc | Old | Total | Fresha | Old | Total | Fresh | Old | Grand Total | |
| Number of scats | 27 | 43 | 70 | 104 | 110 | 214 | 74 | 77 | 154 | 205 | 230 | 438 |
| Number of occurrences | 36 | 54 | 90 | 131 | 124 | 255 | 86 | 85 | 171 | 253 | 263 | 516 |
| Food items | Percent of occurrence | |||||||||||
| Moose, unidentified | 3 | ... | 1 | 12 | 25 | 18 | 9 | 9 | 9 | 9.5 | 14.8 | 12.2 |
| Moose, calf | 39 | 48 | 44 | 43 | 2 | 23 | 60 | 14 | 38 | 48.2 | 15.5 | 31.5 |
| Moose, adult Moose, total | 17 59 | 20 68 | 19 64 | 16 71 |
48 75 | 32 73 | 14 84 | 65 88 | 39 86 | 15.8 73.5 |
47.9 78.3 | 32.1 75.9 |
| Beaver | 17 | 17 | 17 | 12 | 10 | 12 | 7 | 7 | 7 | 11.1 | 10.6 | 10.8 |
| Grass | 8 | 4 | 6 | 9 | 6 | 8 | 3 | 2 | 2 | 3.5 | 2.6 | 3.1 |
| Snowshoe hare | 5 | 6 | 6 | 2 | 3 | 3 | 5 | ... | 2 | 3.5 | 2.6 | 3.1 |
| Soil | 8 | ... | 3 | 3 | tr. | 1 | ... | tr. | 3 | 2 | tr. | 1.7 |
| Unidentified mammal | ... | ... | ... | 2 | tr. | 1 | ... | tr. | 1 | .7 | 1.1 | .8 |
| Unidentified | 3 | ... | 1 | 1 | tr. | tr. | ... | ... | ... | .7 | tr. | 6 |
| Bird | ... | ... | 2 | ... | tr. | tr. | ... | ... | ... | ... | 1.1 | .6 |
| Red fox | ... | ... | ... | ... | tr. | tr. | ... | ... | ... | ... | tr. | tr. |
| Red squirrel | ... | 2 | 1 | ... | ... | ... | ... | ... | ... | ... | tr. | tr. |
| Deer mouse | ... | ... | ... | ... | tr. | tr. | ... | ... | ... | ... | tr. | tr. |
bUnknown age.
cMay through August, plus 3 scats from October.
Although grass or sedge made up about 6 percent of the items, these are not considered food. They often are found in canid scats and even have been reported from mountain lion (Felis concolor) scats (Robinette et al., 1959). Some vegetation may be eaten inadvertently with the prey. Isle Royale wolves eat bloody snow while waiting for a wounded moose to weaken; perhaps in summer they eat blood-spattered grass or even bloody soil. Murie (1944) found that some of the wolf scats containing grass also held several round-worms, and suggested that grass may act as a scour.

Figure 61—All that remains of a
2-1/2-month-old calf, killed by wolves.
Moose remains composed 76 percent of the total (516) items. In scats from May through August, they constituted 74 percent of the occurences, and in "old" scats they formed 78 percent. (Old scats probably were from autumn and winter primarily.) Beaver remains composed approximately 11 percent of the total items, so beavers appear to be the only other important food.
FREQUENCY OF PREDATION
One of the most important figures obtained during this study is the rate of moose kill by the pack of 15 wolves (table 11). (All animals fed upon by wolves are considered "kills," as is discussed on p. 115.) I believe that every kill made by this pack from February 5 to March 4, 1959, and from February 5 to March 20, 1960 was located. In 1961 most of the kills were found, but the wolves' activities were unknown on 11 of the 48 days between February 2 and March 20. During the periods in which the rate of kill was known in 1961, it averaged the same as in 1959 and 1960—one moose per 3 days. However, the wolves once killed two moose in 2 days, and the longest period we found between kills was at least 118 hours, and may have been as much as 137 hours, between March 7 and 12, 1960. The chronological distribution of kills is shown in table 11.
TABLE 11.—CHRONOLOGICAL DISTRIBUTION, AND RATE, OF MOOSE KILL
BY THE PACK OF 15
[Underlined dates indicate known dates of kill; others may vary by a
day]
| 1959 | 1960 | 1961 | |||
| Date | Age and sex | Date | Age and sex | Date | Age and sex |
| Feb. 5 | cow | Feb. 5 | calf | Feb. 2 | cow |
| Feb. 8 | calf | Feb. 7 | calf | Feb. 4 | bull |
| Feb. 11 | calf | Feb. 9 | calf | Feb. 7—8 | calf |
| Feb. 14 | adult | Feb. 12 | cow | (Feb. 9—16 unknown) | |
| Feb. 17 | calf | Feb. 16 | cow | Feb. 17 | bull |
| Feb. 18 | cow | Feb. 20a | bull | Feb. 18 | calfc |
| Feb. 21 | adult | Feb. 22a | bull | Feb. 23 | cow |
| Feb. 24 | cow | Feb. 27 | calf | Feb. 25 | calf |
| Mar. 1—4 | bull | Mar. 1 | calf | Feb. 27 | adult |
| Mar. 5 | calf | Mar. 1—5 | fed on old killsd | ||
| Mar. 7 | calf | (Mar. 6—11 unknown) | |||
| Mar 12a | cow | Mar. 12 | adult | ||
| Mar. 15b | calf | Mar. 14 | calf | ||
| Mar. 17 | calf | Mar. 16 | adult | ||
| Mar. 18—20 | calfc | Mar. 18—20 | bull | ||
| Summation | |||||
| 28 days: 9 moose | 45 days: 15 moose | 37 days: 12 moose | |||
bFound dead, but see p. 137.
c Human activity interfered with previous kill.
dA calf may have been killed during this period.
Apparently, the pack of 15 makes fewer kills than do wolves in other areas. Field men in the Rocky Mountain national parks of Canada determined the rate of kill of two packs of five or six wolves (Cowan, 1947). Each pack killed three elk per 2 weeks, "with indications that two small elk might be taken in a week," Conversion of these figures on a basis of pack size for comparison with those from Isle Royale gives a rate of one kill per 1.6 days. Stenlund (1955) estimated that a pack of 3 wolves in Minnesota would kill about one deer per 4 days, so 15 wolves probably would make one kill per .8 day. Through aerial observations of a pack of 10 Alaskan wolves, Burkholder (1959) found 21 kills (14 caribou and 8 moose calves) made in 35 days, an adjusted rate of one kill per 1.2 days. Some of the differences among these figures can be accounted for on the basis of prey size, as will be apparent in the next section. Undoubtedly, variation in availability of prey and in the methods used to derive these figures also contributes to the differences.
Since Isle Royale's smaller wolf packs never were studied closely for several weeks in a row, little is known about their rate of kill. Probably, both small groups (totaling five) did not together kill over a third the number of moose killed by the large pack. During the 1961 study period (48 days), when I became more proficient in locating kills made by the smaller packs, five of these were found. This is a third of the large pack's expected total (16), although all kills of the small packs may not have been found. These groups probably have more difficulty killing moose than the 15 wolves do, but each carcass should last them longer. Members of the small packs frequently wander far from their kills, so they may hunt new prospects while still able to resort to a previous kill for food. By the time one carcass is eaten, they may have another. In addition, each pack might feed on the other's kill. These speculations are based on limited evidence, but they might indicate direction for future research.
The lone wolf probably kills few moose in winter; it usually feeds on remains left by other wolves. Only once was evidence found that this animal made its own kill. The moose had been wounded and abandoned by the large pack a few days earlier. After finishing it off on March 12, 1961, the lone wolf fed without competition (except from foxes and ravens) at least until March 20.
FOOD CONSUMPTION
No kill was weighed, so all consumption figures are based on moose weights given in the literature. According to information from Kellum (1941), Skuncke (vide Peterson, 1955:77), and Simkin (1962), fully adult cows average about 800 pounds, and bulls, 1,000 pounds (see page 93). Possibly these figures are a bit high for animals killed in winter, but since the amount of possible weight decrease occurring over winter is unknown, these figures will be used. Nine-month-old calves apparently weigh about 300 pounds (modified from Peterson). For ease in assessing kill figures from both the present study and from the literature, I have adapted an arbitrary unit to allow for the varying weights of big game. One "prey unit" is considered to be 100 pounds. Following are the assumed prey units for various ages and species of big game:moose calf, 3; cow, 8; bull, 10; deer, 1; elk, 6; caribou, 3. Adult moose of unknown sex are assumed to be cows, since the sex ratio of kills favored cows strongly.
When the Isle Royale rates of kill are examined in terms of prey units, they do not appear so uniform. The pack of 15 consumed 2.11 p.u. per day in 1959, 1.64 per day in 1960, and 2.21 per day in 1961. Comparable figures calculated from the literature are: Cowan (1947), 3.85 p.u. per day; Stenlund (1955), 1.25; and Burkholder (1959), 2.80. Since Stenlund's figure is an estimate, probably it is not as valid as those from Cowan and Burkholder, which are known rates of kill, or at least minimum rates closely approaching actuality.
Figures of average consumption per wolf-day should include consideration of the weight of uneaten remains. Since remains were not weighed, a standard estimate must suffice. I believe that unconsumed bones, skin, and hair averaged about 50 pounds per adult moose, and 15 pounds per calf. On this basis, the pack of 15 devoured approximately 5,555 pounds of moose in 28 days during 1959, or 13.2 pounds per wolf-day. In 1960, the 16 wolves consumed about 7,000 pounds in 45 days, or 9.7 pounds per wolf per day. The 1961 consumption by 15 wolves was approximately 7,740 pounds in 37 days, or 13.9 pounds per wolf-day.
No average-daily-consumption figures for wolves were found in the literature. However, Wright (1960) reported that African lions (Panthera leo) consumed an estimated .11 to .13 pounds per pound of lion per day, and wild dogs (Lycaon pictus lupinus) ate .15 pounds per pound of dog per day. On the assumption that the Isle Royale wolf pack contains 5 females at 61 pounds, and 10 males at 78 pounds (weights from Stenlund, 1955), the average wolf weighs 72 pounds. Therefore, the average daily consumption rates per pound of wolf per day would be: (1959) .18; (1960) .13; and (1961) .19. These figures compare favorably with Wright's.
Although average rates are useful figures, they also are misleading, for a wolf's feeding schedule is quite erratic. When food is available, wolves gorge; then they may go several days without eating. Young and Goldman (1944:120) explain it as follows:
Of all the members of the canine family, the wolf, when in its prime, can be most irregular in its feeding habits. Equipped with abundant power to kill, the preference is for large prey in order to sustain its large body. When large prey is not available, long intervals between meals may be endured rather than spend time and energy in quest of small animals. Then when opportunity again occurs the animals fill themselves to repletion.
As would be expected, the capacity of a wolf's stomach is extremely large. Several times, the pack of 15 devoured a calf within 24 hours, a rate of about 20 pounds per individual per day. On one occasion these animals consumed approximately half a cow in less than 2 hours. They killed the moose at 2:40 p.m. on February 12, 1960, and immediately began to feed. By 4:10 p.m. only three wolves were feeding, and the carcass appeared at least half eaten. The cow was mature, but possibly it was smaller than average. Even if it weighed only 600 pounds, the 15 wolves ate about 20 pounds apiece in 1-1/2 hours.
Other authors have recorded similar feats. Young and Goldman (1944) reported a wolf stomach weighing 18 pounds and another weighing 19 pounds, 3 ounces. According to Cowan (1947), four wolves devoured most of a doe mule deer in 4 hours; and in 5 days, three wolves consumed two mule deer and a calf elk. Cole (1957) found two instances in which a pack of seven or eight Isle Royale wolves ate about three-quarters of an adult moose in 2 days, a consumption rate of about 35 pounds per wolf-day.
Besides being able to consume great amounts of food in short periods, a wolf also can fast several days with no evident hardship. In 1960, the pack of 15 went at least 95 hours (March 8, 3 p.m. to March 12, 2:30 p.m.) apparently without eating anything except possibly hair and bones which they might have gleaned from old kills. In 1961, half of the large pack spent from March 1 to March 5 without any food except a beaver and scraps from old kills. (Possibly the wolves fed on material cached at old kills, but I think this is unlikely. Before leaving a kill, they clean it so completely that there probably is no extra food to cache.) Young and Goldman (1944) reported on a captive wild wolf which fasted a week and then gorged on the eighth day. E. H. McCleery of Kane, Pa., who has raised wolves—as many as 34 at a time—for many years, wrote me that in winter and spring he feeds his animals every 5 days, and in summer, every 5 to 10 days.
FEEDING ROUTINE
Wolves feed for the first few hours after making a kill. Apparently, a cow moose is not quite large enough for the entire 15 animals to feed on at once, for at the one cow we saw killed, two wolves had to wait off at one side; the others, packed solidly around the carcass, were tugging voraciously at it. After gorging on a fresh kill, the wolves usually curl up nearby and sleep. Each probably feeds at least twice during the first half day, for seldom during this period are there no individuals feeding. About mid-morning the whole pack often heads for an open ridge or stretch of ice, sometimes over a mile away, where each animal sprawls in the sun for several hours (figure 62). A few wander back to the kill now and then if it is not far. Around midafternoon the pack returns to feed. On the second day, if the carcass is large, the pack frequently travels leisurely for a few miles to a resting spot. By then little is left of the kill except the intact skeleton. When the wolves return again, they dismember the skeleton and spend hours gnawing bones. Usually 2, and some times 3, days are spent at the carcass of an adult, and one or 2 at a calf carcass. Although there is much variability in the above-described routine, it seems to be the basic pattern.
Figure 62—Resting attitudes of engorged wolves.
Two calves were examined on the ground soon after being killed, and information was obtained on the parts eaten first. On February 5, 1960, one of a set of twins was killed at 4:40 p.m. By 5:10 p.m. the neck and left side of the chest had been skinned; the heart, part of the lungs, the rump, and the nose were eaten, and there was a hole in the side of the abdomen (figure 63). The second kill, made at 11:45 a.m., March 17, 1960, was investigated within 45 minutes. Most of the meat was missing from one side of the head and throat, and from the upper hind leg and pelvic region; part of one shoulder was eaten. One side of the abdomen was wide open, with intestines pulled out and partly eaten, and the liver was gone. Whether these parts are preferred or whether they merely represent the points of attack is unknown.

Figure 63—Remains of 9-month-old calf 45 minutes after being killed by the large pack.
There is some indication that the pelvic and abdominal regions and the nose are preferred. An adult killed about 7:30 a.m. on August 26, 1960, was examined a few hours later. The wolves had been frightened from the carcass about 8 a.m., and the only meat missing was about 15 pounds from around the pelvis. Undoubtedly, most of the wolves were reluctant to return, for 2 days later just the nose and left side of the abdomen had been eaten. A bull wounded by the large pack and killed by a lone wolf about 6:30 p.m., March 12, 1961, was checked the next day at 11 a.m. The meat and a few pieces of intestine in the pelvic region were all that had been eaten.
Before the wolves abandon a carcass, all the viscera and flesh and about half the skin and hair are consumed. Sometimes the skin is left on the lower legs, but if the carcass is revisited, this is eaten also. Calves killed in winter usually are dismembered completely; all that remain are a chunk of hide, the disarticulated long bones, the mandible and upper tooth rows, and a great patch of hair (figure 64). In summer, the skin and most of the bones of calves apparently are devoured. Cow remains include the skull and anterior half of the backbone in one piece, and the pelvis and posterior part of the backbone in another. The legs are detached from the skeleton, but most of the bones of each remain together. The ends of the ribs and long bones. and the edges of the scapulae and mandibles are ragged from being chewed (figure 65). Usually, bull skeletons are less pulled apart but are thoroughly cleaned of meat. The completeness with which carcasses are consumed may attest that wolves have difficulty obtaining prey.

Figure 64—Remains of the calf in Fig. 63, 24 hours later.

Figure 65—Cow moose remains gathered just after the large pack abandoned them.
Burkholder (1959:9) found the following usage pattern of caribou and calf moose carcasses:
The first parts of the animal eaten are the viscera, except for the stomach contents. The soft parts of the neck and ribs appear to be preferred over the more massive tissue structure. In many cases the entire animal is consumed, including hoofs, long bones, and skull, with only hair and stomach contents remaining.
Parasites and Diseases
Wolves are susceptible to several diseases and physical disorders, and are hosts to many helminth parasites; mange also infests them. Stenlund (1955) has reviewed the literature on the subject. Since his work, Rausch (1958) reported on 43 rabid canids from Alaska, including two wolves. Rausch and Williamson (1959) examined 200 wolf carcasses and listed the helminths found therein.
No wolf carcass was handled during the present study, so little information on parasites and diseases was obtained. However, intermediate stages of two species of helminths, for which the wolf is the definitive host, were found in Isle Royale moose. Taenia hydatigena occurred in two of four adults examined, and Echinococcus granulosus was found in three of them. Adults of both worms have been reported from wolves in several areas (table 12).
TABLE 12.—REPORTED INFECTIONS OF WOLVES BY "ECHINOCOCCUS GRANULOSUS" AND "TAENIA HYDATIGENA"
| Species | Location | Number examined |
Number infected |
Percent infected |
Source |
| Echinococcus granulosus: | Minnesota | 8 | 5 | 63 | Riley (1939) |
| Minnesota | 18 | 5 | 28 | Erickson (1944) | |
| British Columbia | 5 | 1 | 20 | Cowan (1947) | |
| N. Ontario | 58 | 36 | 62 | Sweatman (1952) | |
| Minnesota | 18 | 4 | 22 | Stenlund (1955) | |
| Alaska | 200 | 60 | 30 | Rausch & Williamson (1959) | |
| Ontario | 520 | 103 | 20 | Freeman et al. (1961) | |
| Taemia hydatigena: | Minnesota | 18 | 8 | 44 | Erickson (1944) |
| British Columbia | 5 | 4 | 80 | Cowan (1947) | |
| Minnesota | 18 | 15 | 83 | Stenlund (1955) | |
| N. Ontario | 10 | 10 | 100 | Sweatman & Plummer (1957) | |
| Alaska | 78 | 56 | 72 | Rausch & Williamson (1959) | |
| Ontario | 520 | 39 | 8 | Freeman et al. (1961) | |
An adult Taenia sp. was found by D. L. Allen soon after it had been passed by an animal (presumably a wolf, from the sign). This could have been T. hydatigena, or even T. krabbei, the larvae of which encyst in the muscles of big game. Moose were not examined for this parasite, so it is not known whether the species occurs on Isle Royale. Rausch and Williamson (1959) found the adults in 48 of 78 wolves from Alaska, and Erickson (1944) and Stenlund (1955) reported Taenia sp., which may have been this species, from Minnesota wolves. Peterson (1955) reported that cysticerci occurred in muscles of an Ontario moose.
The effect of these parasites on the wolf is unknown. Erickson (1944) noted that some wolves harbored so many Taenia hydatigena that their intestines appeared blocked. Adults of Echinococcus granulosus are only 2 to 8 mm. long (Chandler, 1955), but according to Rausch (1952:159) heavy infections of Echinococcus granulosus in the final hosts are usual; "in some cases the cestodes cover nearly the entire mucosal surface of the host intestine." According to Monnig (1938:103), "varying degrees of enteritis may be present [in dogs and cats] from a catarrh to a croupous or haemorrhagic enteritis, especially in heavy infections. . . ."
Choquette (1956) experimentally infected dogs with Echinococcus granulosus cysts (as many as seven cysts, from 3 to 10 cm. in diameter, to an individual). He writes (p. 191) that "while it is agreed that dogs can harbor a great number of adult worms without apparent ill effects there are reports of pathogenicity." Haemorrhagic enteritis and rabiform symptoms, diarrhea, asthenia, cachexia, catarrhal inflammation, and death in heavily infected individuals, are listed as effects in dogs. Three of the eight dogs infected by Choquette showed severe diarrhea, weight loss, and asthenia. "Death occurred within a few days of the appearance of symptoms and a month after initial infection. Post-mortem examination showed a severe haemorrhagic enteritis and a very large number of immature worms" (Choquette, 1956:192).
Wolves probably have a greater resistance to these worms than do domestic animals, since the former certainly have evolved with the parasites. Although eight of Choquette's nine experimental dogs became infected, only five passed eggs and segments in their feces, so even all dogs are not equally susceptible. Apparently, the adults of Echinococcus are short-lived. One of Choquette's dogs passed eggs for 8 months and then stopped. Autopsy showed it was worm-free. A second dog, still eliminating eggs, was autopsied 6 months after infection, and senile worms were found. Thus, infections probably do not accumulate for more than 8 months, but in this period, a wolf could become reinfected several times. Wolves probably can support high cestode populations without significant effects, although young, ailing, or old individuals may be affected adversely.

Figure 66—Author checking femur marrow of wolf-killed
moose.
Figure 67—Comparison of normal femur marrow (left) with
fat-depleted marrow.
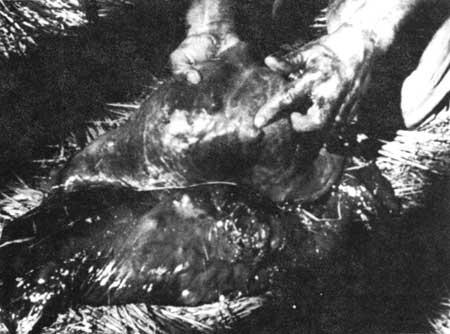
Figure 68—Lung with hydatid cysts from wolf-killed moose. Photo by D. L. Allen.

Figure 69—Hydatid cysts from wolf-killed moose. Photo by D. L. Allen.
Possible Causes of Population Stability
According to Young and Goldman (1944), the average wolf litter contains seven pups. In Minnesota, Stenlund (1955) found that 8 litters averaged 6.4 young. The mean size of four litters from British Columbia was five (Cowan, 1947). This information alone might lead one to suppose that wolves are prolific and that wolf populations have a high rate of turnover. However, facts do not agree with this supposition.
Cowan found that in the Rocky Mountain national parks in Canada, three packs which had been checked carefully showed no significant increase during two breeding seasons. In northern Alberta, Fuller and Novakowski (1955) poisoned 3 entire packs and found an age ratio of 3 pups to 10 adults. On Isle Royale, two wolf packs have each remained the same size for three winters; indeed, the pack of three apparently has failed to increase since at least early 1957, when Cole reported on it. The apparent inconsistency between these data and the fact that wolf litters are large probably results from one or more of the following factors: unproductive animals, prenatal losses, mortality factors, and emigration and immigration.
UNPRODUCTIVE ANIMALS
There appear to be at least four categories of unproductive members of wolf packs: (1) surplus males, (2) immature animals, (3) senile individuals, and (4) social subordinates. Murie (1944) found three males and two females, all at least 2 years old, at one den in Mount McKinley National Park; two more adult males joined this pack in late summer. Fuller and Novakowski found one to three extra adults in five out of six packs which they poisoned.
Young and Goldman (1944) quoted a report that the sex ratio was equal in a catch of 68 wolves from New Mexico, and Fuller and Novakowski found an even sex ratio in 58 poisoned wolves in Wood Buffalo National Park, Canada. However, males comprised 15 of 25 wolves shot in British Columbia (Cowan, 1947), and 100 of 156 animals taken in Minnesota (Stenlund, 1955).
An attempt was made to sex Isle Royale wolves on the basis of size and mating behavior. Stenlund found that, in a sample of 114 wolves, males averaged 17 pounds more than females, and Fuller and Novakowski (1955) reported an average difference of 13 pounds, although in each study, weights overlapped between the sexes. On Isle Royale, only one member of the large pack consistently appeared small, but in photographs, three and possibly four smaller animals are evident (figure 57). During all the mating activity observed, I never noticed more than three individuals being pursued amorously at one time. Perhaps other females in estrus were not noticed, or possibly some were not in estrus. Nevertheless, since three or four animals were interested in each of the three "known" females, I believe there is a substantial preponderance of males in this pack. One member of the pack of two, and one of the pack of three, also are smaller and may be females.
Since males mature in 3 years, and females in 2 (Young and Goldman, 1944), most wolf populations will contain a number of unproductive young. In an increasing population, or in one with a high turnover rate, immature animals could compose a high percentage. The only wolves on Isle Royale which were thought to have been immature were the three lanky animals in the large pack.
Wolf populations which are not hunted or trapped heavily probably include a substantial number of old animals. Young and Goldman report that old age for a wolf is 10 to 14 years. Such old wolves, according to these authors, often travel alone and subsist on old kills and carrion; presumably these would be senile. Fuller and Novakowski examined a very old male which had small testicular volume and no demonstrable spermatogenesis. There is at least one lone wolf on Isle Royale which probably is in this category, and several of the other animals also could be senile.
The most important class of unproductive animals might be the social subordinates. Mykytowycz (1960) found that in a captive population of wild European rabbits (Oryctolagus cuniculus) the dominant females bred much more effectively than the subordinates. Retzlaff (vide Christian, 1958:477) also noticed this phenomenon in a population of laboratory mice (Mus musculus). One might gather from Schenkel (1948:fig. 50a) that the same applies to wolves, for he uses the phrase "suppressed, but not entirely 'frigid' females." It would seem advantageous for the alpha individuals in a society to be the most effective reproducers, for probably they are physically the best, or at least the most aggressive. Thus, the non-reproductive members of a group could help supply the food to the reproductive members and their young. Murie (1944) believed that one of the packs he studied was organized in this manner. An extra female even helped care for the young, and stayed with them one night when the mother went hunting.
Laboratory studies of mice by Davis and Christian (1957) have shown a correlation between social status and weight of the adrenal glands, the lowest-ranking individuals (i.e., the most stressed) having the heaviest glands. A similar study by Christian (1956) demonstrated that the animals with heaviest adrenals reproduced least effectively. The same principle might apply to wild wolves, although this would be difficult to prove. It might operate both within a pack, inhibiting the reproductive ability of low-ranking members, and between packs, causing reproductive inhibition in repressed packs. In discussing territoriality, Elton (1950) stated that species having few effective natural enemies tend to be self-regulatory, and Murie (1944) and Stenlund (1955) agreed that territoriality would tend to control wolf numbers. The manner in which this might operate is unknown, but perhaps the above-mentioned relationships are involved.
PRE-NATAL LOSSES
Animals from populations of high densities generally have lower reproductive rates than those in less-dense populations. Hoffman (1958) showed an inverse relationship between density and ovulation rate in Microtus montanus. Davis (1949) was able to increase pregnancy rates of brown rats (Rattus norvegicus) in Baltimore by reducing the population.
This inverse relationship between density and litter size might becaused by variations in nutrition and/or in social stress. Mason (1939) discussed the effect of nutrition on reproduction. An increase in amount and variety of nutrients just before the breeding season causes a higher ovulation rate in sheep (Clark, 1934; Stoddard and Smith, 1943; Belschner, 1951). Cheatum and Severinghaus (1950) and Longhurst et al. (1952) agree that ovulation rate in deer also is affected by the level of nutrition. According to Frank (1957), the litter size of the vole (Microtus arvalis) depends particularly on the quantity and quality of food. Lack (1946) asserted that when avian predators dependent on mice face a food shortage, they fail to breed; when mice and lemmings are excessively common, these birds may raise two broods and have clutches twice the usual size.
Stevenson-Hamilton (1937:257) writes of a similar relationship in African lions:
It was discovered that lions . . . could barely be kept static in numbers. So easy was it for them to catch their prey, that a lioness was accustomed to produce cubs at about twice the normal rate; in place of the usual two or three, she brought forth as many as four or five in a litter; while of these, instead of one or two only, probably all, or nearly all, were able to survive to maturity.
Whereas the effect of nutrition on reproduction has been studied for many years, the role of high-density stress is just beginning to be evaluated. Most of the significant work has been done with laboratory mice. By studying populations of mice given unlimited food and water but varying in number per cage, Christian (1956) discovered that high population density suppressed reproduction in both sexes. One of the manifestations of reproductive suppression was decreased litter size. In a later experiment, Christian and Lemunyan (1958) found that all 10 of their crowded females had bred, but that 7 of these lost all their embryos through both pre- and post-implantation mortality.
Of course, it usually is extremely difficult to separate the roles of nutrition and of social stress in wild populations of high density. However, Kalela (vide Christian, 1958:491) found that a wild population of the redback vole (Clethrionomys rufocanus) increased in density to a peak and then suddenly collapsed, despite the abundance of available food. Christian et al. (1960), studying a die-off in a herd of sika deer (Cervus nippon), ruled out malnutrition as a cause of mass mortality, and concluded that high-density stress was primarily responsible.
If high-density stress sometimes controls wild populations, conceivably this factor at least contributes to the stability of predator populations, especially in territorial species. In this connection, it may be significant that Isle Royale has one of the highest wolf densities reported (table 6).
MORTALITY FACTORS
Mortality might take one of several forms, but probably it occurs most frequently in the pup class. Cowan (1947) reported on a bitch which apparently had lost an entire litter of young. Fuller and Novakowski (1955) found a ratio of 9 pups to 36 adults in autumn and concluded that this indicated a pup-mortality rate of about 90 percent within the first 6 months. This estimate apparently is based on the questionable assumption that each pair of wolves produces six young each year. Nevertheless, the ratio found by these authors does suggest a high death rate among pups.
There are several possible causes of pup mortality. Conceivably, the bitch might obtain enough food to produce a full litter, and then because of seasonal changes, possible pack break-up, or other adverse circumstances, she might not get sufficient food to nourish all her young. This might apply especially to Isle Royale wolves. Pups should be born there in the third or fourth week of April—about the time the ice goes out. Moose then can take refuge in the water so perhaps are less vulnerable at this time. New calves, which composed much of the wolves' summer diet, are not born until mid-May.
According to Speelman (1939), domestic dog pups require two or three times as much food as adults of the same weight. This should apply to wolf pups as well, so any food shortage could be crucial for them. Stevenson-Hamilton (1937) and Wright (1960) observed behavior in East African lions, which, if duplicated by wolves, would be disastrous to pups. The females and cubs feed on kills only after the males have satiated themselves, and frequently there is little left for the cubs.
Figure 70—Large pack crossing ice.
Young and Goldman (1944) report that often one or two whelps are more aggressive than the others. Probably if a food shortage existed, these individuals would steal all the food, leading to the eventual death of their littermates. Lee Smits of Detroit, Mich., who has raised wolves, suggested another idea to me. He believes that, during the violent activity which occurs when wolves are fed, one pup might bite another, become excited, and end up devouring it too. In this way, only the most vigorous individuals would survive.
In a special supplement to the Kane (Pennsylvania) Republican paying tribute to Dr. E. H. McCleery and the population of captive wolves he has kept for 40 years, McCleery claims that "if a mother lobo has one outstanding pup she may keep that one—eat the others. Also she will eat an injured pup." Although these observations pertain to captive animals, it is possible that under stress, even a wild wolf would act this way.
Social stress during the period that young are being raised could be an important factor. A laboratory experiment with crowded mice produced the following conclusion (Christian and Lemunyan, 1958:517) :
. . . suppressed growth of progeny nurtured by crowded mothers, persisting for at least 2 generations, was due to quantitatively and/or qualitatively deficient lactation resulting from crowding. Such attenuation of the effects of crowding may explain the long-continued decline in natural populations following peak levels and a precipitous crash in numbers.
Again, interaction of the nutrition and stress factors probably would be more important that the action of either alone.
No information is available on the incidence of pup mortality from injuries by prey, but it might be quite significant, particularly on Isle Royale. Even the large pack of adult wolves must chase many moose before killing one, and the animals obviously are afraid of a threatening moose. MacFarlane (1905) and Stanwell-Fletcher (1942) reported instances in which a wolf was found badly injured by blows from a moose. Naive and inexperienced puppies might not respect moose as adults do, and in the excitement of a chase might be especially vulnerable to the deft kicks of their intended prey. Even experienced adults might perish in this manner, although apparently this has not happened on Isle Royale during the study.
The diseases and parasites discussed previously might be important in controlling wolf numbers in certain locations. It is doubtful that these are directly significant on Isle Royale, because no evidence of adult mortality was found. If pathogenic organisms were primarily responsible for pup mortality, they probably would cause death to a few adults, too. Indirectly, such parasites as Taenia hydatigena and Echinococcus granulosus in heavy infections might add to any general stress affecting the wolves, and, therefore, might contribute to whatever reproductive inhibitions there may be.
Old age eventually may be a significant mortality factor on Isle Royale. Seton (1937) writes of a male wolf in the National Zoological Park, which lived over 16 years. If any of the original Isle Royale immigrants remains, it would be at least 14 years old. Wolves over 10 years of age generally are considered old; they usually have worn and broken teeth and find it difficult to obtain food (Young and Goldman, 1944). Such individuals visit old carcasses more often than other wolves. At least one Isle Royale animal, the lone wolf, spent much time at old kills. Since this individual was not fully accepted by the pack, it probably was inferior in some respect, perhaps in age.
EMIGRATION AND IMMIGRATION
Because wolves did immigrate to Isle Royale, the possibility exists that other movements to or from the island have occurred since, or will occur. Significant immigration probably depends upon the following: (1) a high wolf population or a food shortage on the nearby mainland, causing "pressure" for animals to seek new territory; (2) a solid, snow-covered "ice bridge" to the island; (3) the type of reception given the newcomers by the residents; and (4) the ability of the immigrants to kill moose. Since wolves apparently did not populate Isle Royale during this century until about 1949, it seems reasonable to assume that the necessary combination of circumstances does not occur often.
Continuous ice does not connect Isle Royale with the mainland every year, so the ice-bridge factor probably is most critical in determining whether movement in either direction occurs. Cole (1957) collected several reports of years when ice connected the island with the mainland. In two of the three winters during the present study, extensive sheets of thin, drifting ice spanned the lake to Canada several times. Reports, apparently originating from pilots of high-flying aircraft, once claimed that the entire lake was frozen over.
However, after each high wind, the ice piled onto the north shore of Isle Royale, leaving the lake open. Thus, reports of ice bridges to the island should be viewed cautiously. Nevertheless, from February 15 until at least March 21, 1961, the ice with stood several severe winds, and appeared safe for even a vehicle to cross.
Besides depending on suitable ice, movements of wolves from Isle Royale also probably would depend on the animals having a strong reason to leave their home range, or at least a great desire to travel (perhaps only temporarily) to new territory. We did witness one apparent attempt by the large pack to leave the island. On March 1, 1960, the 16 wolves were heading northeastward in Rock Harbor at 2:25 p.m., after traveling about 29 miles from their last kill. They reached Blake's Point, the northeast tip of the island, at 4:40 p.m., and by 4:45 were about a mile due north of the point.
The wolves continued toward Canada another half-mile, gradually curved eastward, and then headed toward Passage Island. The ice was smooth in places, but elsewhere it consisted of older chunks frozen together with new ice. The animals were cautious about crossing from one type of ice to another. Although most of the wolves appeared reluctant to proceed, the leader seemed determined. Several times this animal returned to the hesitant pack and apparently tried to urge the members on. They continued about one-half a mile until encountering ice composed of many small, sharp pieces frozen together. After testing this, the pack returned to Isle Royale (figure 53).
I do not believe the wolves were heading to Passage Island, for they could have taken a more direct route. They might have been able to reach Canada, since there had been little wind the previous week. However, they probably would not have returned, even if they had wanted to, for a few days later the wind had shifted the ice, leaving large cracks.
During the 1961 study period, when a substantial ice bridge existed, the pack certainly could have emigrated, but no sign of an attempt was observed. Nevertheless, it always is conceivable that someday one of the packs might wander off and never return. Future investigators should attempt to watch the wolves very closely during periods when continuous ice extends to Canada.
Moose Herd
Isle Royale moose probably are intergrades of the eastern subspecies Alces alces americana and the northwestern subspecies A. a. andersoni (Peterson, 1955:6). The only available weights of wild Isle Royale moose are a few supplied by Murie (1934). However, Kellum (1941:5) kept six Isle Royale moose and their offspring in corrals on the Michigan mainland from 1936 to 1941, weighed them each month, and obtained the following information:
Newborn calves weigh from 25 to 35 pounds, and gain from one to two pounds daily for the first month, and from three to five pounds daily the next month. Males weigh more than females at similar ages. A year old male may weigh from 400 to 600 pounds; at two years about 700 pounds; at three years about 900; and from then on his weight will vary with the seasons, from 900 to 1,200 pounds. . . . Females follow similar trends, weighing about 400 pounds when one year old, 600 pounds at two years, and from 600 to 800 pounds during maturity, depending on the condition of the animal and the season.
These weights may be high, for the moose were fed maximally and had little room for exercise.

Figure 71—Mature bull moose eating aquatic plants in early June.
Skuncke (vide Peterson, 1955:77) produced a growth curve for European moose (Alces alces alces) based on weights of 637 animals from Sweden. This shows that cows with calves achieve maximum weight, about 750 pounds, when 5 years old; cows without calves average about 850. Bulls weigh about 900 pounds when 5 years old and approximately 1,050 pounds in their 11th year. This subspecies supposedly is slightly smaller than most North American subspecies.
 Figure 72—Moose feeding in Washington Harbor. |
 Figure 73—Moose track. |
From the data of these authors and from Simkin (1962), it seems probable that adult Isle Royale cows average about 800 pounds, and bulls, 1,000.
The food of moose varies with the season; Peterson (1955) discussed the general food habits of the species.
On Isle Royale, moose depend primarily on aspen, white birch, balsam fir, and mountain ash for winter food. Aldous and Krefting (1946), who made an extensive and detailed analysis of the island's winter moose food, listed 28 browse species. Summer food consists of leaves and twigs of many of these species, plus various forbs and aquatics. Hazelnut, large-leaved aster, thimbleberry, pondweeds, and water lilies are among the favorite summer foods. The aquatics are sought from early May to late August, especially by bulls. Murie (1934) presented an annotated list of the island's summer moose foods, and Brown (n.d.) discussed the gross changes in vegetation wrought by the high moose population.
Incidental observations made on moose pelage change indicated that the summer coat first became apparent about mid-June and that by mid-July, all animals had new pelage. Calves had winter coats by the end of August.
Great variation was noticed in the degree of antler development among various bulls. Antler formation began several weeks earlier in bulls with large antlers, presumably older individuals. Bulls with large palms were observed about the same time as animals at least 2 years old were noted with nothing but antler pedicels. The following observations provide a general idea of the amount of variation:
| May 15 | spike horns. |
| May 15 | palmate antlers about 18 inches. |
| May 16 | antler pedicel only. |
| May 22 | antler pedicel only. |
| June 17 | buttons. |
| June 21 | spikes about 4 inches long. |
| June 22 | large palmate antlers fully formed except for small tines. |
| June 25 | spikes about 4 inches long. |
| June 28 | small palm forming. |
| July 2 | spikes about 4 inches long. |
| July 19 | very large palm with outer tines formed but blunt. |
| Aug. 23 | large palm complete and some velvet gone. |
Antlers had been shed by most bulls by early February when the winter studies began. However, a few animals were seen with cervina-type antlers during February, and the latest we noticed a bull with antlers was March 12.
NUMBERS AND DISTRIBUTION
The first extensive aerial census of moose on Isle Royale was made by Aldous (Aldous and Krefting, 1946) in February 1945, with a Waco five-seated biplane. Eight parallel strips were flown, and a 30 percent coverage of the island was obtained; 122 moose were seen. Allowing for an arbitrary 20 percent error, the workers estimated the size of the population at 510. Another count in 1947, reported by Krefting (1951), produced an estimate of 600 animals. Cole (1957) made the next aerial census, but he attempted a complete count. A Piper Cub was used to fly narrow strips at 400 to 500 feet altitude until moose or tracks were located. Often only one of a group of animals was spotted, so the pilot spiraled the aircraft downward to about 100 feet, and the running moose were counted. Cole observed 242 animals, and tracks of another 48, and estimated the population at 300. He believed that in favorable weather 90 percent of the island's moose could be counted by this method.
Trotter (1958) in Ontario also was impressed with the results of an intensive-search method of survey. He compared results of three types of censuses made on the same areas: (1) intensive survey by helicopter, (2) transects with a Beaver aircraft, and (3) intensive search with the Beaver. He concluded (p. 6) that ". . . the intensive search method [with Beaver] produced uniformly high counts of moose, whereas the transect method count was low and not consistent enough."
During the present study, Cole's method was used. The island was divided into convenient-sized plots with natural boundaries, a technique suggested by Trotter. With the 90-horsepower Aeronca Champ traveling 70 to 80 m.p.h. at 400 to 500 feet altitude, we flew strips paralleling the length of the island. Strips varied in width with terrain and cover but approximated one-eighth of a mile, and they overlapped to insure complete coverage. Duplicate counts of the same moose usually were avoided easily because strip length was short (3 to 6 miles) and locations of previously counted animals could be remembered from one strip to the next.

Figure 75—Cow moose with new summer coat.
Since I counted moose each day only after observing the wolves, censusing took several days. Each night the possibility existed that moose from a censused area would wander into an uncensused area and vice versa. Because no reason was found for animals to travel consistently only in or out of a censused area, I assumed that these movements would compensate for one another.
Each time we observed a moose, even in an open area, we "buzzed" it at about 100 feet, causing it and any nearby unseen individuals to run and be counted. (Lack of fresh snow rendered tracking useless.) This procedure is time-consuming, but results in a much higher count. Banfield et al. (1955) also found that frequently moose are not seen from the air until frightened from their beds.
 Figure 74—Aeronca Champ—used throughout the study for following and counting wolves and moose. |
The first census was attempted in 1959, from March 8 to 13. Because of a low gasoline supply, I counted only two-thirds of the island; 176 moose were seen. Eighteen animals had been recorded on the uncensused third incidental to wolf observations, so there was a minimum of 194 moose on the island. This figure undoubtedly was low.
In 1960, many more moose were seen incidental to other work than had been observed the previous year, probably because of better weather. A complete census was taken between February 13 and March 2, involving 45 hours of flying on 10 days, and 529 moose were seen. On the area censused in 1959, 439 moose were found in 1960. The difference (263) between counts during the 2 years obviously cannot be attributed entirely to reproduction. Although other factors may be involved, I believe that the most significant cause of the disparity is variation in weather conditions, especially in wind velocity. Banfield et al. (1955:521) stressed that "unfavorable weather conditions such as strong winds and snowstorms may force big game to take shelter in forests and thus become harder to see from the air." In 1959 strong winds frequently forced us to discontinue censusing, whereas 1960 was characterized by clear and calm days. Observations in 1960 showed that on windy days significantly fewer moose frequented openings.
The most important effect of wind variation on moose censuses should be emphasized, for, apparently, much greater differences in counts can becaused by this factor than would usually be caused by variations in population. For instance, if Cole's census was hindered by strong winds, his count could have been much too low. Even the 1960 census could below. However, I do not believe that a much higher count will be achieved unless the population does increase, for apparently optimum conditions prevailed during that census.
Any other factor, such as time of day, which may influence the location of moose in reference to conifer cover may affect a count profoundly. Even under the best conditions, many moose undoubtedly are missed. Once when one moose was spotted, circling and diving eventually revealed six other animals were with it; if the one had not been seen, seven would have been missed. Often the pilot sighted moose that I overlooked, but he could not observe much since he had to keep the plane on course. Because many moose are missed during aerial censuses, some authors suggest that a compensatory figure be applied to the results. By comparing aerial and ground censuses on the same area, Edwards (1954) decided that aerial-census figures would be more accurate if increased by 22 percent. Banfield et al. (1955) agreed that aerial estimates are about 20 percent low.

Figure 77—Moose and tracks in winter, as seen during
moose census.
 Figure 76—White birch killed by moose many years ago. |
Peterson (1955) reported that on St. Ignace Island, where conifer cover is dominant, only about a third of the moose present could be counted from the air. During a ground census in Montana, Knowlton (1960) observed 53 moose; in the same area, he counted only 15 from an aircraft 8 days later.
Because of these factors, I believe that a conservative estimate of the number of moose on Isle Royale in March 1960, was 600. The approximate density then is 3 per square mile. Peterson (1955:202) summarized reported population data from Minnesota, Nova Scotia, Newfoundland, and Ontario, and concluded the following:
In summary, it appears that in eastern North America an average density of 1 moose per 5 square miles might be regarded as a "normal" average density for a great portion of the range. One moose per square mile is probably a relatively high density under most conditions, while 2 or more moose per square mile represent an approach to maximum carrying capacity for most large regions. While much higher densities undoubtedly occur in restricted areas, at least temporarily, they have not been observed on large areas (1,000 square miles or more) in eastern North America on a sustained basis.
Moose densities in western North America are not directly comparable to those in the east because, in the former area, summer and winter ranges frequently are miles apart, and reported winter densities are only temporary. In general, western densities are much higher than densities in eastern regions (Peterson, 1955). For instance, Spencer and Chatelain (1953) estimated that the best winter range in south central Alaska (willow, birch, aspen, and cottonwood) can support 5 to 10 moose per square mile "under proper use."
Moose inhabit all of Isle Royale and most of the surrounding islands. Some areas have higher densities than others, however, and local variations are evident between summer and winter distributions. The burns (figure 7) and swamps seem to have the highest concentrations during all seasons, but especially in winter (figure 13). The paucity of animals on the northeast third of the island in winter probably is caused mostly by a shortage of browse, although lack of extensive swamps also may be a factor.
According to Peterson (1955), snow usually does not confine moose to swamps at it does deer, but moose do seem to use swamps for protection from wind. Tracks show that many moose headquarter in swamps and venture from these each day for food.
In spring there is a notable shift of moose to the northeast third of the island. Evidently the attraction there is aquatic plants which thrive in the many beaver floodings in the area. (Terrestrial herbs do not appear on Isle Royale until late May, but apparently aquatics begin to grow by early May.) I have seen 8 bulls and yearlings in Ojibway Lake at one time and 12 other moose with in half a mile of the lake. It is unusual to spend a few hours in this location in May or June and not see moose. Use of marshy areas decreases throughout July and August and ends almost completely in September. Peterson (1955) noticed a similar usage pattern of lakes and streams by moose in Ontario.
REPRODUCTION
Accurate knowledge of the sex ratio in any moose population is difficult to obtain, for the sexes differ considerably in behavior, habits, and distribution. Schierbeck (1929) in Nova Scotia found a ratio of 2,232 bulls to 6,175 cows, and Spencer and Chatelain (1953) reported a bull-cow ratio of 38 to 62 for 5,319 Alaskan moose. However, Pimlott (1953) and Peterson (1955) found even sex ratios in large samples from Newfoundland and Ontario, respectively. Conversely, Pimlott (1959a) stated that kill data from Newfoundland, Sweden, and Norway, and fetal and observational data from Newfoundland show a preponderance of males. Nevertheless, he admitted (p. 447) that ". . . a reasonable doubt exists that the sex ratio of the population actually departs from 50:50."

Figure 78—Mature bull in June.

Figure 79—Cow with 2- to 3-week-old calf.
In Montana, Knowlton (1960) found a summer ratio of 100 cows to 206 bulls (based on 248 identifications), whereas the observed winter ratio for 104 animals was 100 cows to 131 bulls. The sex ratio of hunter-killed moose in the same area was 15 cows to 16 bulls, and of 27 calves in winter was 100 females to 92 males. Knowlton discussed possible reasons for observation of an unbalanced sex ratio when such actually does not exist.
Because of the errors inherent in the method, no attempt was made to determine, on the basis of ground observations, the sex ratio of the Isle Royale moose population. Such information was obtained by a method probably less subject to bias. An aerial survey was made during 11 hours between October 27 and 31, 1959. The same technique was employed as described for the winter moose census, although a 90-horse power Piper Cub on floats was used, piloted by Jack Burgess of Tower, Minn. D. L. Allen and I alternated as observers. Coverage included the entire area northeast of a northwest tangent to the southwest end of Hatchet Lake (approximately 40 percent of the island), and 150 moose were seen. Of these, 33 were calves, 57 were bulls, and 60 were antlerless. Of the antlerless, 32 definitely were cows, since they were accompanied by calves; the other probably also were females, for according to information by Peterson 1955:90) and Cringan (1955:240—246) yearling bulls normally have antlers. However, in Montana, Knowlton (1960) observed animals identified as yearlings, with only 3/4 to 1-1/2-inch buttons as late as September 26, so possibly some apparently antlerless moose on Isle Royale actually had antlers. Nevertheless, because we obtained close aerial views of most animals, and because we did observe spikes and other small antlers on 14 moose, I believe that this type of error was small, and that the census indicated a balanced sex ratio.
According to Murie (1934), the rutting season on Isle Royale extends from mid-September to mid-October, with its height in late September. These dates coincide in general with those furnished by Peterson (1955) for the rutting season in Ontario.
Murie saw the first calf of a season on May 28 and believed that the peak of calving occurred in late May and early June. Information from the present study corroborates this. Cooperators reported the following earliest dates for first calves seen: May 26, 1959; May 20, 1960; May 19, 1961.

Figure 80—One of twin fetuses removed from a moose found dead
April 24, 1959.

Figure 81—Young bull swimming across Rock Harbor, July
1960.
The last observation was by W. Leslie Robinette, of the U.S. Bureau of Sport Fisheries and Wildlife, who judged the calf to be a few hours old. Peterson reported that Ontario moose also calve in late May and early June.
The possibility of an unusually early birth on Isle Royale was indicated by the size of twin fetuses removed from a moose found dead on April 24, 1959, and examined on May 8. The male weighed 17-1/4 pounds and was 29 inches long, and the female weighed 16 pounds and was 28-1/2; inches long. Both had open eyes and were completely covered with hair; their incisors had erupted but were still soft (figure 80). No weights of newborn or fetal twins were found in the literature, but probably a twin would weigh a few pounds less than a single calf. Murie (1934) examined a fetus on May 20 which weighed 22 pounds and was 36 inches long. The smallest of four healthy newborn calves in captivity in Michigan weighed 24 pounds, although a fifth, which died when 2 days old, weighed only 13 pounds (Kellum, 1941).
An attempt was made to determine the approximate number of calves produced annually by the moose population. Observation forms were distributed to Park Service personnel, commercial fishermen, and other summer residents. These people were asked to record every moose sighting, even if they thought they saw the same animals every day. Since several cooperators seldom left their own sections of the island, many of the reports probably involved relatively few animals. In 1959, 291 observations were made by 14 cooperators (including the investigator) after May 26, when the first calf was seen, and calves composed 25 percent of them. A critical statistical evaluation of this type of sampling cannot be made, for the number of different animals observed is unknown. If 100 different individuals were observed, the 95 percent confidence limits would be .17 and .33. In 1960, 20 cooperators reported 359 observations, of which calves composed 15 percent. The 95 percent confidence limits would be .10 and .20, if 150 different animals were seen.
All biases in this method would tend to decrease the calf percentage. A higher proportion of yearlings probably is observed than exists in the population, for these recently independent individuals lack the caution of more mature animals and probably also wander more. Secondly, a cow often ventures afield without her calf, especially before the calf is very old (Peterson, 1955). During the remainder of the summer, young moose frequently stray from their parents far enough to be missed by an observer. Another source of bias might be the summer concentrations of bulls, for when an observation is made, several animals may be involved. The latter bias would be important only when relatively few cooperators are reporting, such as during this study.
Other authors (Peterson, 1955; Pimlott, 1959b) also have concluded that surveys of this type indicate lower percentages of calves than actually are present. In Montana, Knowlton (1960) found a ratio of 100 cows to 69 calves (based on 137 observations) in summer, and 100 to 78 (80 observations) in winter, suggesting the unreliability of summer figures. On Isle Royale, Murie (1934) encountered a similar trend. In summer, he found 26 percent of 103 cows followed by calves, whereas in autumn, calves accompanied 40 percent of 42 cows, and in spring, 46 percent of 83 cows. Although there are several possible explanations for these unexpected trends (including small sample sizes), the most likely seems to be a summer bias against calves. In Newfoundland, Pimlott 1959b:399) found that "the percent age of cows observed with calves (35) was approximately half the percent age of pregnant cows (73), the difference being caused by observational biases Therefore, calf-total population ratios obtained in summer during the present study must be considered minimum.
The only such ratio obtained in autumn resulted from the aerial survey made in late October 1959. Of the 150 moose seen, 33, or 22 percent, were calves. (The 95 percent confidence limits are .16 and .28.)
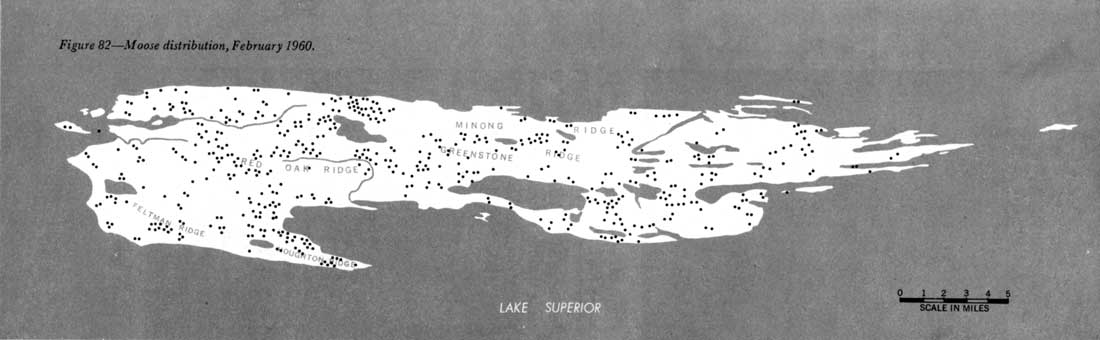
Figure 82.—Moose distribution, February 1960.
(click on image for an enlargement in a new window)
Reported twinning rates for North American moose populations vary from 2 percent to 28 percent, on the basis of field observations (Pimlott, 1959b), although some local herds might not include any twins (Knowlton, 1960). Four of the eight rates considered by Pimlott were between 11 percent and 18 percent. On Isle Royale, 20 (38 percent) of 53 cows seen with calves in the summer of 1959 were accompanied by twins. If only 25 different cows were seen, the 95 percent confidence limits would be 19 percent and 57 percent. If the lower rate is assumed, the number of calves per cow-with-calf is 1.19. (Also see page 170.)
The most significant moose-population statistic is the yearling-total population ratio. As will be shown later, if an Isle Royale moose survives its first year, chances are excellent that it will live several more. Thus, application of this ratio to the estimate of total population size provides an estimate of annual recruitment. The few yearling-total population ratios which have been obtained for Isle Royale are shown in table 13.
TABLE 13.—YEARLING-TOTAL POPULATION RATIOS REPORTED FOR ISLE ROYALE
| Year | Months | Sample size |
Percent of yearlings in populationa |
Source |
| 1930 | May-June | b128 | c21.0 | Murie (1934) |
| 1953 | Feb.-Mar. | 66 | 20.0 | Hakala (1953) |
| 1957 | Feb. | 252 | 15-23.0 | Cole (1957) |
| 1959 | Mar. | 176 | 17.0 | Present study |
| 1960 | Feb. | 529 | 17.0 | Present study |
| 1961 | Feb.-Mar. | 133 | 10.5 | Present study |
aExcluding newborn calves.
bCows and yearlings only.
cCalculated from Murie's cow-calf ratio, with assumption that sex ratio was equal.
During aerial censuses in late winter, short-yearlings (=calves) some times are difficult to distinguish. However, Cole (1957) noted from previous groundwork with Isle Royale moose that when a cow and calf are spooked, they flee together; pairs of adult moose spilt up. Using this information during his aerial survey, he attempted to classify short-yearlings. He believes that possibly 58 (23 percent) of the 252 animals observed were in this category, but his conservative estimate was 15 percent. During the present study, an attempt was made each winter to sample the yearling-total population ratio. In 1959 a low fuel supply prohibited extensive circling and diving, so not all moose observed were aged. Among the 176 animals seen, there were 52 pairs: 18 cows with calves, 18 pairs of 2 adults, and 16 undetermined pairs. In addition, two cows with twins were seen. If it can be assumed that half of the undetermined pairs were cows with calves, then 17 percent of the sample was composed of calves (95 percent confidence limits are .12 and .22). In 1960, 89 (17 percent) of 529 moose observed were calves, and since this sample probably was almost a total count, the 95 percent confidence lim its are .16 and .18.

Figure 83—Calf swimming between islands in mid-July. Photo by B. A. Mech.
Only 133 animals were sampled in 1961, and calves composed 10.5 percent (95 percent confidence limits are .06 and .15). Apparently, this was an exceptionally poor year, for the ratio is the lowest reported from Isle Royale. Since the previous summer's sample indicated that calves composed only 15 percent of the population (compared with 25 percent for 1959), perhaps the calf crop was small.
The average yearling-population ratio probably is about 17 percent, because this is the mean of all the reported figures and because it constituted three of the seven estimates (table 13).
PARASITES AND DISEASES
Unfortunately, adequate information concerning the general health of Isle Royale moose was not obtained. Since Isle Royale is a National Park, wildlife is protected by law, so no hunter-killed carcasses were available for examination; and no animals could be collected during the present study. In all, only six relatively intact carcasses were examined: two adults which died as a result of accidents, two wolf-killed adults, and two wolf-killed calves. Only the lungs, livers, and hearts of five of these were searched for parasites, but the stomachs and intestines of the six also were inspected. In addition, the bones of 48 other (wolf-killed) moose were examined. Although information from these sources applies primarily to wolf-killed moose, it was established that certain parasites and diseases occur in the Isle Royale herd.
The Winter Tick. The following excellent account of the life history and significance of the winter tick (Dermacentor albipictus) is provided by Cowan (1951:42):
This is a one-host tick. The newly hatched seeds become active with the first autumn frosts, climb the vegetation and, with front legs widespread and waving, wait a passing large mammal. Once on a host they feed three times with appropriate periods of rest and two molts. Except in the [West] Coastal area the tick remains on its host all winter long. Although it is at first so small as to escape detection, by early spring the distended, blood-filled bodies of the adults are conspicuous and frequently reported.
The female lays about 4,000 eggs that hatch in six weeks or so but the young remain sluggish and clumped together until the autumn frosts waken them to a winter of blood sucking. It is one of the most serious parasites of big game mammals. It is frequently present in vast numbers. The writer once estimated 7,200 ticks to be present on a mule deer that was at point of death from tick attack at Devona, Alberta, and has seen moose and elk with many more than that. The ticks congregate on the ears, along the lower throat and chest, and on the shoulders, rump and tail region and flanks but may occur anywhere on the body.
This species is particularly damaging because of the period of its activity when the game animals are having their worst time of the year with food shortages and severe weather. Young animals are most subject to attack and suffer more severely. Individual calf moose that have been watched through the winter appeared to be in excellent condition in the autumn, became weaker, thin, and began to rub the hair from parts of the body by late December, suffered progressive weakness, partial paralysis, and death in February or March. Older animals are not immune and James Hatter has well documented accounts of many moose in the Cariboo region perishing from tick attack. In March and April, when the ticks are dropping off, the wounds left bleed freely and the animal's trail is spotted with blood, when it shakes itself the snow over several feet is pink with blood spatters, and every bed is blood-soaked. The stronger animals recover, the weaker die.
Additional life history details are furnished by Cameron and Fulton (1927).
It has not been proved that ticks are a primary cause of moose mortality, although many weak, emaciated, or dead animals have been found heavily infested (Cameron and Fulton, 1927; Lamson, 1941; Olsen and Fenstermacher, 1942; Peil, 1942; Hatter, 1950a; Peterson, 1955). Olsen and Fenstermacher selected mostly ailing and abnormally acting moose to examine, and these may have been infested secondarily; a third of the 36 moose examined harbored no ticks, whereas others harbored thousands. Hatter (1950b) found that high moose mortality in British Columbia resulted from a tick-malnutrition complex, and Ritcey and Edwards (1958) concluded that ticks alone do not seriously weaken moose. Murie (1951) believes that the many dead elk which he found heavily tick-infested were parasitized secondarily, but he did suspect ticks of killing one young moose. Whether ticks are a primary or secondary factor, they do affect moose populations significantly.
Hickie (1936) reported heavy tick infestations on Isle Royale moose, but found that wild-trapped individuals kept in corrals showed no ill effects from the parasites. However since most animals which he found dead of malnutrition were heavily parasitized, possibly ticks were partly responsible for their deaths. Although Cole (1956) did not mention finding these parasites on Isle Royale moose, he noticed that several animals had lost considerable hair in February and March, a good indication of tick infestation.
Only three intact carcasses could be examined for ticks during the present study. A calf killed by wolves in mid-March 1960, had a "moderate" infestation. A 4-year-old bull, which died as a result of an accident in late January 1961, harbored approximately 2 ticks per square inch on about half its body, but a wolf-killed bull (probably at least 13 years old) was infested with approximately 10 ticks per square inch in several places on his rump. The hides of most wolf-kills were torn and scattered, so the degree of infestation could not be determined from these. However, all seven of those that could be checked for ticks in 1961 harbored them. The hide of one very old individual was relatively intact; a high tick population (14 per square inch in places) was present. Lamson (1941) reported a density of 12 ticks per square inch on 116 square inches of a Maine moose.
Because of irritation from these ectoparasites, infested animals rub against trees (Fenstermacher and Jellison, 1933). The hair comes off easily where ticks are numerous (Wallace, 1934), and the neck and flanks usually are denuded first. Naked areas are noticed on many Isle Royale moose by late February. Between mid-May and early June 1960 and 1961, large bare areas were evident on all of 65 animals observed plainly. These denuded areas may have resulted from shedding, which occurs at this time (Peterson, 1955). However, since so many moose in February and March are in this condition, and since most moose show such large naked areas weeks before new hair is apparent, I believe the condition results from tick infestations (figure 84). If that is so, most, if not all, Isle Royale moose are parasitized by ticks to some extent.

Figure 84—A moose lacking most of its hair in
May. This may have resulted from a heavy tick infestation the previous
winter.
Hydatid Cysts. These cysts contain the larvae of the tapeworm Echinococcus granulosus, which in habits the intestine of the wolf and other carnivores. Eggs and mature proglottids pass out with the feces and into water or onto vegetation. The intermediate host, which may be a moose or any of several other big-game species, ingests the eggs while eating or drinking. Schiller (1954) demonstrated that mice could be infected experimentally by blowflies (Phormia regina) fed on infected feces, so this method of transmission also may be important. After an egg hatches in the digestive system of the herbivore, the larva enters the blood stream, circulates, and eventually encysts in a lung (usually). The cyst grows and, after about 5 months, reaches one centimeter in diameter (Faust, 1949); numerous brood capsules containing more larvae begin to form (Monnig, 1938). Cysts frequently reach golfball size in moose, but Chandler (1955) reports one containing 10 to 15 quarts of fluid (host not mentioned). The life cycle is not complete until the cyst is eaten by an appropriate carnivore, in which the larvae become adults.
Apparently, the parasite is widely distributed, for it has been reported from Minnesota (Olsen and Fenstermacher, 1942), Ontario (de Vos and Allin, 1949), Manitoba (Hadwen, 1933), Saskatchewan (Harper et al., 1955), Alberta (Cowan, 1948), British Columbia (Cowan, 1947), Northwest Territories (Banfield, 1954), and Alaska (Rausch, 1959). Peterson (1955) found that Echinococcus was the most common parasite encountered in Ontario moose. The reported incidences of infection in moose vary between 30 percent and 68 percent (table 14).
TABLE 14.—REPORTED INCIDENCES OF INFECTION OF MOOSE WITH HYDATID CYSTS ("ECHONOCOCCUS GRANULOSUS")
| Location | Number of animals |
Percent infected |
Source |
| Minnesota | 33 | 36 | Olsen & Fenstermacher (1942) |
| Ontario | 29 | 60 | Sweatman (1952) |
| Alaska | 11 | 36 | Spencer & Chatelain (1953) |
| Saskatchewan | 96 | 30 | Harper et al. (1955) |
| British Columbia | 34 | 68 | Ritcey & Edwards (1958) |
| Alaska | a78 | 31 | Rausch (1959) |
Hydatid cysts may affect animals seriously, for they sometimes occur in large numbers. Although Ritcey and Edwards (1958) found a mean of 7.7 and a mode of 1 cyst in the lungs of 23 infected moose, 1 harbored 32. They provided (p. 143) the following description of the effects of heavy infection:
Another cow, on March 6, 1955, was trapped in a large corral for tagging. As men approached the trap she advanced with defiant behavior, stopped, began to tremble violently, then sank slowly to the ground breathing heavily. Breathing became weaker, and the animal was dead in four minutes. Autopsy revealed she was about four years old. She had the heaviest hydatid infection that we have encountered. There were three hydatid cysts in the liver and at least 30 in the lungs, ranging from 1/4 to 2 inches in diameter. The cow was pregnant with twin calves, had heavy fat deposits on the omnentum, and fat 3/4—1 inch thick over the kidneys. The only unusual feature found, aside from cysts, was an excessive amount of fluid in the pericardial sac.
Other sick, abnormally acting, or weak moose with heavy hydatid infections were reported by Law and Kennedy (1933), de Vos and Allin (1949), and Peterson (1955). Cowan (1948) examined an elk lung which was replaced almost completely with cysts, and Fenstermacher (1937) found 50 cysts in the lungs of one moose, and 250, replacing about three-quarters of the lung, in another.
Figure 85.—Life cycle of the hydatid tapeworm,
Echinoccus granulosus: A. Eggs passed with feces of wolf. B. Moose
eats eggs with browse. C. Larvae circulate to lungs and encyst. D. Larvae
reproduce asexually in cyst. E. Wolves kill moose and inadvertently eat
cysts. F. Larvae mature into adult tapeworms and live in wolf's intestine.
The first record of hydatid cysts from Isle Royale was reported by Sweatman (1952:481); he stated that "five of eight moose were found infected in 1933 on Isle Royale by Dr. D. Coburn." During the present study, three of the four adult moose examined harbored these parasites. The most heavily infected was a cow about 8 years old with 57 cysts (5 to 20 mm. in diameter) in her lungs. Because such a heavy infection was found and because wolves have been devouring moose for several years on Isle Royale (and therefore propagating the worm), it seems probable that the parasite infects a majority of the island moose.
There is more than one possible explanation for the present occurrence of Echinococcus on Isle Royale. Coyotes or foxes, which probably spread the parasite before 1933, might have continued to do so. Although probably neither of the smaller canids killed moose undoubtedly both fed on carcasses. Cowan (1948) suspected coyotes of maintaining hydatid tapeworms in Alberta. According to Riley (1939), foxes are primary hosts of the parasite in Europe, although he knew of no records of fox infection in the United States. Another possibility is that wolves visited Isle Royale occasionally and spread enough eggs to propagate the species. However, the most likely explanation is that the progenitors of the present wolf population harbored adults of Echinococcus when they arrived.
Taenia hydatigena. Cysts of this species usually are found in the livers of various big-game animals. After a cyst is eaten by a wolf or other suitable carnivore, the larva develops into a tapeworm which dwells in the intestine of this primary host. If the eggs, which are passed with the carnivore's feces, are eaten by a moose or other ungulate, they hatch in the intestine, and the larvae migrate to the liver where they encyst.
Cowan (1951) reported that in Alberta and British Columbia the incidence of infection with this parasite is high, but that seldom are there over 12 cysts per animal. In Minnesota, the opposite was found. There were 75 cysts in one moose (Fenstermacher, 1937), but only 5.8 percent of 34 moose were infected (Olsen and Fenstermacher, 1942). Sweatman and Plummer (1957) reported that 15 of 17 moose from Ontario harbored Taenia hydatigena cysts. Of 32 moose autopsied in British Columbia, 84 percent were infected (Ritcey and Edwards, 1958). The only comment on their effect was that they cause no apparent harm (Cowan, 1951).
No previous record was found of this parasite on Isle Royale, but during the present study, Taenia hydatigena cysts were discovered in two of four moose livers examined. One contained 5, the other 10. These specimens were identified by W. W. Becklund of the U.S. Department of Agriculture and were deposited in the U.S. National Museum Helminthological Collection as 57208 and 57210.
Jaw Necrosis. Of 91 moose mandibles and/or upper jaws from Isle Royale, 13 (14 percent) were swollen, porous, and abscessed (figure 86). No attempt was made to isolate a causative organism, but the symptoms are similar to those described for "lumpy jaw," or actinomycosis (Monlux and Davis, 1956). According to these authors, "an anaerobic microorganism, Actinomyces bovis, which is more closely related to fungi than to true bacteria, produces these lesions involving the jaws." The disease infects either or both jaws, and since many of the moose remains found had upper or lower jaws missing, the incidence of this disease probably is higher than the figures indicate. In 1929 and 1930, of the 20 remains of Isle Royale moose, 11 (55 percent) showed similar symptoms (Murie, 1934).
Figure 86—Necrotic moose mandibles.
The causative organism requires a laceration or abrasion through which to enter the jaw. Most infected specimens collected during the present study were from old moose, and the abscess usually centered around the first molar. This molar is the oldest and, therefore, the first to wear below the gum line. When this occurs, apparently the occluding upper molar wears into the gum, permitting entry of the pathogen. While attempting to age moose by tooth wear, Passmore et al. (1955:233) found that "necrotic lesions appeared to have developed very readily when excessive wear had reduced the height of any tooth below that of the normal gum line." Ritcey and Edwards (1958) reported that 5 of 34 autopsied moose had actinomycosis, and that it was most severe in old animals.
Murie (1944:117—120) discussed actinomycosis and related disorders in detail. He believes that the disease is chronic and that heavy infections can be debilitating. Certainly the abscesses and misshapen jaws and teeth resulting from actinomycosis would impair proper mastication, which might be serious to herbivores.
Lungworm (Dictyocaulus). This nematode infects the bronchial passages of moose and other big game, and its larvae are coughed up and out, or are swallowed and passed with the feces. They eventually climb low vegetation to await ingestion by a new host (Cowan, 1951). Olsen and Fenstermacher (1942) discovered Dictyocaulus viviparus in 42 percent of 33 sickly moose from Minnesota, and Lamson (1941) reported D. hadweni (considered by some workers as a synonym for D. viviparus) from a Maine moose. Heavy infections can be debilitating, as the following from Cowan (1951:53) shows:
In addition to the obstructing effect of the adult worms in the lungs, the irritation induced frequently causes a thickening in the walls of the air passages that is characteristic. The inflammation induced and the interference with normal lung function often provides the opportunity for bacterial invasion and a broncho-pneumonia results. This causes the animal to have a husky cough and difficulty in breathing: diarrhoea is usual. Even if death does not result directly from the lung-worm attack the animals are so weakened that they are vulnerable to winter conditions or predator harassment. Animals usually die after a prolonged decline and show the same marrow symptoms of fat loss accompanying other debilitating diseases.
During the present study, Dictyocaulus sp. was found accidentally in one moose. While counting Echinococcus cysts in the excised lungs of an animal examined on August 26, 1960, I noticed a live lungworm. It was identified by M. B. Chitwood of the U.S. Department of Agriculture as Dictyocaulus sp. and was placed in the U.S. National Museum Helminthological Collection as 56879. No carcasses were examined for this parasite, and since it has not been reported previously from Isle Royale, no information is available on incidence of infection or significance to the island herd.
Other Parasites and Diseases. In summer, most moose on Isle Royale are pestered by swarms of flies, and there are raw spots near the hocks on at least a few animals. Murie (1934) observed similar lesions on Isle Royale moose. Peterson (1955) reports the condition from Ontario animals, and believes that it probably results from the collective efforts of the insects. Both authors identified specimens of the pests as moose flies (Lyperosiops alcis). Whether the deer-flies (Chrysops) and black flies (Simulium), abundant on Isle Royale, also parasitize moose is unknown. Undoubtedly, flies are a great nuisance, but the amount of harm they cause has not been determined.
Since complete pathological examinations of moose were not attempted during the present study, there probably are other parasites and/or diseases not yet discovered in the Isle Royale herd. Peterson (1955) summarized the information available on moose parasites and diseases, and discussed approximately 25 species.
MORTALITY
In areas without wolf populations, moose probably succumb to a variety of factors such as malnutrition, old age, diseases, and accidents. Most of these factors cause a gradual decline, so in areas with high wolf populations, dying animals probably are eliminated by predation, and mortality caused directly by any other factors should below. Such seems to be the case on Isle Royale. Since the 15 to 16 wolves alone eat (and probably kill) an average of 1 moose per 3 days in winter, presumably few moose get a chance to die directly of causes other than predation.
Information on moose mortality was obtained from investigation of as many carcasses and old remains as could be located. These were discovered by two methods. In winter, the aircraft was used to track wolves to carcasses which they fed upon. Some moose were seen killed, and tracks showed that wolves killed several of the other animals which they ate. In many cases, the network of wolf tracks around a "kill" prevented positive determination that the wolves had dispatched the moose. However, except in two cases, no reason was found to indicate that they had not, and chances are good that even the two excepted moose were killed by wolves. Therefore all remains found fed upon in winter will be considered kills; any error resulting from this assumption undoubtedly is small. Fifty-six such kills were found, 51 of which were investigated on the ground. Although most of these were taken by the large pack, several eaten by the smaller packs are included.
The second method used to locate moose remains was ground search in spring and summer. Reports from alerted Park Service personnel and other field men facilitated this work. (W. L. Robinette and L. W. Krefting, of the U.S. Bureau of Sport Fisheries and Wildlife, provided information from 18 remains they examined during a 3-week browse survey in 1961.) A total of 72 remains are included in this "random" sample, and these should represent year-round mortality from most sources and over a period of several years.
Accidents constitute the only moose mortality factor that is relatively unrelated to predation. Only three accidental moose deaths came to my attention during this study. A cow about 5 years old was found on a rocky shore near Rock Harbor Lodge on April 24, 1959, by a construction crew. This animal had heavy kidney, heart, and omental fat and her femur marrow was normal. No wolf wounds were found, but small patches of hair were sheared from her head and legs, and the four right posterior ribs were broken. The cow may have fallen through ice and drowned. Peterson (1955) believes that this type of mortality is especially important in early winter and spring.
The carcass of a month-old calf washed up on Scoville Point some time before July 25, 1959, when it was reported; no sign of wolf attack was found. Possibly the animal drowned. Peterson (1955:193) reported that "drowning seems to be an important factor in calf losses Murie (1934) found remains of two Isle Royale moose which probably had drowned.
The third known moose death for which wolves were not responsible occurred on January 31, 1961. A 4-year-old bull had tangled his antlers and neck in an extension cord near a building at Windigo. He spent several days there, becoming increasingly entangled. Park Service personnel were eventually forced to kill the bull.
Murie (1934) found remains of 6 animals that had been mired or had caught a foot in roots, but no indications of this type of mortality were found during the present study.
Wolf predation undoubtedly is the most significant moose mortality factor on Isle Royale. A possible measure of its importance is afforded by data from randomly discovered moose remains. Such remains frequently can be judged "eaten by wolves," or "not eaten." (Characteristic signs of wolf feeding are: widely scattered bones; separation of the vertebral column into two pieces; and obvious chewing on the edges of the scapulae, the posterior-ventral angles of the mandibular rami, and the ends of long bones. Chewing is evident even after the bones have lain for several years.) Remains of 47 moose were found which could be judged "eaten" or "not eaten." Of these, 34 (72 percent) had been chewed by wolves. Most of those showing no wolf work were well weathered, and some may have dated from the "pre-wolf" period.
Of course, wolf-chewed moose bones do not mean necessarily that wolves killed the moose; Isle Royale wolves do eat carrion. On June 23, 1959, I investigated the putrid carcass of a bull lying a few feet offshore in Chickenbone Lake. Fully formed antlers without velvet indicated that the moose had perished in autumn or early winter. Wolves recently had detached two legs and eaten them on shore. By July 7, the entire carcass had been dragged ashore and eaten.
However, apparently the wolves sometimes wound a moose and leave it. (The bull mentioned above might have been such a victim.) A wounded animal might wander far before dying, and in summer might decompose before the wolves discover it. The bones would not appear chewed, even though wolves were responsible for the animal's death. Data from such remains would tend to compensate for data from remains of an animal that the wolves had eaten but not killed.
Wolf-Moose Relationships
The welfare of a wolf population is related intimately to the availability of prey. In many areas wolves prey on two or more species, and if one becomes relatively unavailable, another provides sustenance. From most reports it appears that an adult moose is one of the most formidable prey animals in North America, so when another species is available, wolves tend to depend heavily on the other species. Peterson (1955:175) reported that in the St. Ignace Island area, "where moose were much more abundant than white-tailed deer," moose remains composed only 36 percent of 76 wolf scats collected, where as deer remains comprised 57 percent. In Alaska, Burkholder (1959) established that 14 of 22 wolf kills were caribou and 8 were moose. He believes that these species were killed in proportion to their availability. Nevertheless, six of the seven ageable moose were calves, and the other was a yearling, whereas several of the caribou were adults. In Mount McKinley National Park, Murie (1944:57) concluded that although caribou, Dall sheep, and moose are available, ". . . moose are not readily taken by wolves." Cowan (1947) found that, in the Rocky Mountain national parks of Canada, where several big-game species are present, moose remains occurred in 9 percent of 420 wolf scats, and most of the remains were those of calves.
MacFarlane (1905) and Stanwell-Fletcher (1942) each reported on a wolf which had been injured badly in an encounter with an adult moose, so it is evident why wolves are cautious in dealing with these animals.
Isle Royale wolves must resort to moose for their primary food, for no other big game is present. Because of this, the resident packs undoubtedly are expert at hunting and killing moose—any inept individuals probably perished long ago. Aerial observations of 66 hunts provide the basis for the following analysis of the wolves' hunting techniques. The number of hunts observed and hours of observation each year are summarized in table 15. Since the wolves hunt as they travel, the figures are based on observations of the wolves traveling, exclusive of rest periods and time spent on large bays or lakes. Moose often occur in groups, so the number of moose involved is given in each case. All observations included in the table were of the large pack or part of it.
TABLE 15.—OBSERVATIONS OF WOLVESa HUNTING
| Year | Hours of observation |
Number of hunts |
Number of moose involvedb |
| 1959 1960 1961 |
9 35 24 |
6 33 27 |
15 66 51 |
| Total | 68 | 66 | 132 |
bSince moose often are in groups, one hunt usually involved more than one animal.
When observing wolves hunting, we habitually flew ahead of them and spotted the next moose along their trail. This allowed observation of both wolf and moose behavior before, during, and after attacks. By refueling when the pack rested or passed through areas devoid of moose, we minimized the possibility of having to leave in the middle of a hunt. The fuel cache at Mott Island plus a 5-gallon can of fuel carried in the aircraft were advantageous in this respect. Each hunt witnessed is described in the appendix.
HUNTING HABITS OF THE WOLVES
After the wolves leave a kill, apparently any moose encountered is subject to attack. Once the animals showed interest in a moose only 35 minutes after leaving a kill. In an other case, they killed a moose within 4 to 10 hours after they abandoned their last carcass; and eight times they dispatched a moose within 26 hours after leaving a previous victim. They did almost all of their killing within 48 hours after they left their previous prey. In Alaska, a pack left a moose carcass at noon one day, and by the next morning had dispatched and eaten another moose (Burkholder, 1959). The average distances between kills made on Isle Royale are given in table 8.
The Isle Royale wolves employ a method of hunting moose which differs from methods reportedly used for other species. In Ontario, Dunne (1939) learned that in searching swamps for deer, packs split into twos and threes. Cowan (1947) and Stenlund (1955) found that wolves use a "line abreast" formation upon reaching areas to be hunted. When the predators hunt points and islands for deer in Minnesota, part of the pack drives while a few animals wait on the ice to intercept any prey flushed (Stenlund, 1955). Wolves hunt elk in the Canadian Rockies by traveling single file on ridges and rushing any quarry below them (Cowan, 1947). According to Murie (1944), the Mount McKinley wolves hunt Dall sheep by coursing through the hills hoping to surprise an animal at a disadvantage; when hunting caribou, they merely approach one of the many herds and begin chasing it.
Isle Royale wolves apparently do not have special places to hunt. Everywhere they travel is hunting ground, although more moose are killed in some areas than in others (figure 87). Areas of high moose-kill are characterized by dense moose populations and proximity to well-traveled wolf routes, as is evident by a comparison of figure 87 with figures 82 and 47.
Figure 87.—Distribution of kills.
(click on image for an enlargement in a new window)
The wolves' most common method of hunting is to travel regular routes single file until they scent a moose (figure 88). Most of the moose detected were within 300 yards upwind of the wolves. However, in one open region, the predators apparently sensed a cow and two calves about 1-1/2 miles away. After getting the weak scent of such distant moose, the pack travels toward them until it locates them more precisely.

Figure 88—The large pack hunting.
In certain cases the wolves detected moose 125 yards downwind or 200 yards crosswind. However, I once saw a moose browse undiscovered for 20 minutes, 100 yards downwind of the resting pack. In several instances wolves seemed to scent moose downwind or crosswind, but apparently could not locate them and eventually left. Commonly, when the wolves sense a moose, all stop and "point." Each stands stiffly with nose upwind and ears alert for 10 to 15 seconds, probably verifying the exact location of the moose. Often the animals assemble closely, sniff noses, and wag tails before starting toward the prey.
The wolves did not sense all the moose judged to be within range of detection. Of 160 animals in 85 groups which appeared to be within range, 29 (19 groups) went undiscovered. Some of these were within 150 yards, whereas others were as far as half a mile away. Moose were judged within range if circumstances appeared similar to those in which others had been detected during the many observed hunts. Apparently, topography, cover, local wind direction, and previous behavior of the moose all influence its "detectability."
A less-used hunting technique is tracking. When the wolves cross a fresh moose track, they follow it, single file. Sometimes a couple of animals follow downwind of the track parallel to the others but several yards ahead. On one occasion the wolves scented fresh tracks of a moose that had been working up a small valley below them. They did not actually come upon the tracks but were able to follow them from the ridge about 25 yards above. As the tracks got fresher, most of the wolves headed into the valley and began tracking. One paralleled them on the ridge and finally located the moose, which had moved up the side of the valley. An other time the wolves scented moose sign 20 minutes old from about 50 yards upwind, after the moose had left the vicinity.
From the aircraft, I could determine that a moose trail was fresh only when the moose which made it was nearby. Thus, the wolves frequently may have passed up fresh tracks without my knowledge. On February 6, 1961, the pack of 15 crossed tracks 1 minute old, but only 1 wolf followed them. It gave up after 25 yards and returned to the pack. Before and after this occurrence the wolves were chasing moose, so their unconcern apparently did not result from a lack of motivation.
After detecting a moose, the wolves head excitedly toward it single file, but they check their speed until the moose bolts. If the moose makes a stand, the wolves lunge at it from all sides, trying to force it to run. They are readily frightened by its charges, however, and seldom get close (figure 89). When a moose charges, the wolves scurry several yards away, tails between their legs. If they cannot make the moose flee, they leave. They may decide to depart within half a minute, but sometimes they harass a moose intermittently for 5 minutes before leaving. I once saw a pack harry a standing moose for 3 minutes, finally force it to run, and then kill it within about 10 minutes.

Figure 89a—Fifteen wolves approach a moose in burn area.
Figure 89b—The moose stands its ground.
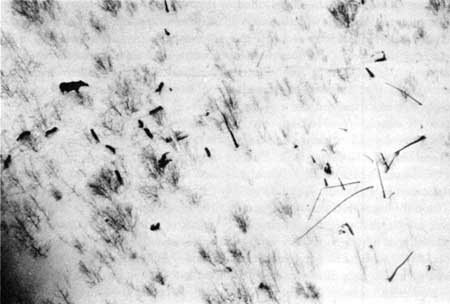
Figure 89c—Wolves harass the moose from a distance.

Figure 89d—The moose holds off wolves; after 5 minutes, wolves leave.
If a moose runs at the approach of wolves, the predators suddenly spring forward with great bounds. Their gait appears exhausting, but they can maintain it for at least 20 minutes. After long runs, they rest for at least 10 minutes. During our observations, the wolves ran faster than the moose through snow less than 2 feet deep, and sometimes within 200 or 300 yards they overtook animals which had a 150-yard lead. However, on one occasion the pack took about a one-quarter of a mile to catch up to a moose with a 100-yard head start. Burkholder (1959) reported an incident in which nine wolves covered 300 yards through snow 2 to 3 feet deep while their prey, a yearling moose, ran only 100. In my experience, pursued moose do not always run at top speed; they seem to depend more on endurance.
Young and Goldman (1944) reported that a wolf was clocked at 27 m.p.h. for 200 yards, but Minnesota wardens chased a wolf on a frozen lake for 4 miles, at 35 to 40 m.p.h. (Stenlund, 1955). The latter rate correlates well with the maximum speed reported for moose (35 m.p.h., by Cottam and Williams, 1943). Thick swamps, heavy cover, blowdown, or snowdrifts slowed the wolves but did not hinder moose. Apparently wolves are aware of their limited ability under adverse conditions, for they sometimes fail to chase nearby animals which run through snowdrifts or blowdown. Hatter (1950a) found that in British Columbia "wolves cannot prey successfully on moose in deep soft snow."
The wolves abandoned 20 observed pursuits without catching up to their intended victims. In these cases, either the moose had too great a lead, or adverse running conditions hampered the wolves. Sometimes one or two wolves got within a few yards of a moose and gave up if the rest of the pack was far behind. Usually if a moose maintained a 100-yard lead for 10 or 15 seconds, the wolves discontinued the chase unless they were gaining. Crisler (1958:106), studying wolves and caribou in Alaska, also was impressed with ". . . how quickly the wolves had judged when a chase was useless."
During extended pursuits, most of the wolves follow single file in the trail of the moose until they over take the animal, undoubtedly making travel easier. Frequently some wolves try shortcutting the moose, but if it turns, these individuals may head in the wrong direction and lose the pack.
After the wolves overtake a moose, most of them remain strung out behind (figure 89), but some stay alongside, apparently awaiting the opportunity for attack. Since pursued moose sometimes travel 2 or 3 miles through several types of cover and over varying terrain without being assailed, probably the physical condition of the moose determines its fate. Although I distinguished no behavioral difference between the three adults seen attacked and the several which fled unharmed, probably the wolves discerned a difference. Two of the moose were killed within 100 yards of where the wolves first encountered them. (The other was wounded within 100 yards but then was abandoned.)
It seems likely that wolves can detect any weakness or inferiority from the behavior of a moose. Certainly if an animal is not strong enough to outlast its pursuers, it will be killed, and this presumably is why the wolves chased some individuals so far without attacking them. Perhaps the two animals killed were debilitated enough so that the wolves immediately detected this and did not hesitate to attack. Two reports in the literature are pertinent here. In Wood Buffalo National Park, Fuller (1960) watched at least 10 wolves approach to within 25 feet of 4 bison. The only bison that showed concern was a wounded one; the others continued ruminating. On two other occasions, Fuller saw (from the air) wolves within a herd of bison which paid them no attention. In East Africa, Wright (1960) watched wild dogs pass near several groups of gazelles (Gazella thomsonii) without frightening them. Only one individual became panic-stricken and ran—it was chased and killed. In these instances, even humans detected behavioral differences between healthy or confident" animals and insecure ones.
Extensive observations of wolves hunting caribou in Alaska show that a primary technique is to chase caribou long distances until a weak or inferior individual is located (Murie, 1944; Crisler, 1956). Murie saw tracks of a chase that lasted 3 or 4 miles, and Crisler witnessed a 5-mile chase. Dall sheep may be pursued for one-half mile over rugged terrain (Murie, 1944), and even deer sometimes are chased vainly for long distances (Dunne, 1939). Thus, it appears that the long-chase technique is employed by wolves in many areas and that it probably serves as a test to distinguish vulnerable individuals.
The technique that wolves use when pursuing a cow moose and calf is to attempt to separate them. While some animals harass the cow, others remain beside the calf, and as soon as the cow charges a wolf, those guarding the calf close in. If the cow fails to keep up with the calf, if thick cover causes the two to separate, or if the cow becomes too involved with chasing off wolves, some animals immediately assail the calf. If the others keep the cow occupied for half a minute, the calf probably is doomed, for two wolves can handle a 9-month-old calf easily. It seems amazing that any cow and calf could survive such strategy. However, eight instances were observed in which cows and calves ran from wolves, and only in three of these were calves killed. This further attests to the profound respect the wolves have for the hoofs of a moose.
Since an insufficient number of hunts by smaller packs and lone wolves were observed, generalizations cannot be made regarding the hunting habits of these wolves. The reader is referred to Hunting Account 31 in the appendix, which involves the pack of three, and 42, 43, 70, and 71, concerning a lone wolf. Although no evidence was obtained that a single wolf can kill an adult moose (unless wounded), the fact that an individual was observed seriously attempting this indicates that at least the wolf thought it might succeed. Young and Goldman (1944) stated that one wolf can kill a full-grown moose, and Cowan (1947:159) reported that in the Rocky Mountain national parks of Canada, "several instances of single wolves killing moose and elk were noted."
ACCOUNTS OF WOLF ATTACKS
Information on the killing techniques of the wolf was obtained from eight hunts in which moose were killed or wounded. Complete, successful hunts of three calves and one adult were witnessed, plus parts of three hunts in which adults were killed, and one in which an adult was wounded and abandoned. In addition, certain information was obtained from an adult killed in summer. The following accounts (numbered for their chronological position in the hunting accounts included in the appendix) are edited versions of field notes. Because of the distances, speeds, and number of moose and wolves involved in each account, and because of the necessity for constant observation from the rapidly moving aircraft, notes are incomplete in some accounts. All distances are estimated.
2. (February 24, 1959. About three-quarters of a mile northeast of Siskiwit Lake Outlet, and about three-eighths of a mile inland.) At 6 p.m., 10 of the 15 wolves were traveling along the shore of Siskiwit Lake about 1 mile ahead of the others. Suddenly they stopped, and several pointed more or less crosswind for a few seconds toward three adult moose three-eighths of a mile away. Heading inland single file to an old beaver meadow, they traveled downwind a few hundred yards, veered, and continued for 250 yards until directly downwind of the moose. Then they ran straight toward the animals, which were still browsing when the wolves were within 150 yards. Two of the moose sensed the wolves 25 yards away and began running. The wolves gave chase a few yards until they spotted the third moose, which was closer and had not left. They immediately ran the 50 feet to this animal and surrounded it.
A few seconds later the moose bolted and the wolves followed in its trail (figure 90). Soon five or six animals were biting at its hind legs, back, and flanks. The moose continued on, dragging the wolves until it fell. In a few seconds the animal was up, but it fell a second time. Arising again, the moose ran through the open second-growth cover to a small stand of spruce and aspen, while the wolves continued their attacks; one wolf grabbed the quarry by the nose. Reaching the stand of trees, the moose stood, bleeding from the throat, but the wolves would not attack.

Figure 90—Wolves pursuing a moose near Malone Bay.
Within a few minutes most of the wolves were lying down, including the last five, which had caught up. Two or three continued to harass the moose without actually biting it, and the moose retaliated by kicking with its hind feet. Whenever the animal faced the wolves, they scattered. Although the moose was bleeding from the throat, it appeared strong and "confident." At 6:30 p.m. we left because of darkness.
The next morning at 11:15 a.m. the wolves were gone. The moose lay within 25 feet of where it had made the stand. After we made several low passes, it finally arose and moved on. Although walking stiffly and favoring its left front leg, the moose was not bleeding and seemed in good shape. The wolves were 16 miles away feeding on a new kill.
3. (March 1, 1959. About a quarter-mile south of Lake Desor, and about half a mile southwest of the northeast end of the lake.) From 10:10 a.m. to 5:05 p.m. the 15 wolves lay on Lake Desor, but at 5:05 they began traveling along the shore. Several seemed to point inland, but mating activity obscured this somewhat. After searching the vicinity and finding no moose nearby, we headed for Mott Island to refuel.
When we returned at 6 p.m. the wolves had a bull (as determined later) surrounded in a small stand of hardwoods. He was bleeding steadily from the throat, and had difficultly holding his head up. About 150 square feet of the surrounding snow was covered with blood. The animal's lower left hind leg was bloody, and he leaned against a tree, keeping his right hind leg centered under him. (Chances are good that this animal had been assailed the night before and then temporarily abandoned.)
Most of the wolves were yards away, resting and playing, but a few were licking the bloody snow. One wolf in particular, whose legs were covered with blood, was harassing the moose. It stayed near the bull most of the time, often nipping at the injured leg. However, each time the moose faced it or any nearby wolves, they scrambled away. At 6:30 p.m. we left because of darkness.
Unfavorable weather prevented a check on the situation until March 4. At 10:45 a.m. the bones of the moose were scattered around the spot where we had seen him last. The wolves had just left and were 10 miles away. An examination of the remains showed this to be a bull in wear-class VI (Passmore et al., 1955).
7. (February 5, 1960. About 200 yards south of Siskiwit Lake and 1 mile west of Wood Lake.) At 3:50 p.m. we left the 16 wolves heading across Siskiwit Lake near Ryan Island. After refueling, we tracked them to the southeast shore of the lake, up the first ridge, and along it northeastward. At 4:35 p.m. we saw them running upwind on the open ridge toward a cow and two calves about three-quarters of a mile away. The way the wolves had veered upon reaching the ridge about 1-1/2 miles from the moose suggests they had smelled the animals at that distance.
When still three-quarters of a mile from them, several of the wolves stood on a 100-foot ridge and pointed toward the moose, which now faced them. The first few animals charged off the ridge and ran toward the moose but a little north of them. Two wolves were far ahead, and two others ran south of the trail left by the moose.
The cow and calves eventually headed toward Wood Lake, but upon encountering a steep drop-off, they turned southward. The first two wolves sped after the moose, gained rapidly, and overtook them within a quarter mile. As the moose ran through open second-growth birch, one wolf remained on each side.
The cow was immediately behind the calves, and twice she feinted toward the wolves, which leaped out of the way. Most of the pack began catching up, and as the moose entered a small cedar swamp (the nearest conifer cover), four or five animals tore at the rump and sides of a calf and clung to it. Within 50 feet, the calf went down in a thick clump of cedars. The cow and the other calf continued through the cover with two wolves still following for 20 yards. When these wolves gave up, the moose stopped and returned 50 yards toward the wounded calf. Gradually, however, the moose drifted back toward where they originally had started. Most of the wolves concentrated on the wounded calf, which remained where it had fallen. The cedars obscured our vision, but the calf appeared dead within 5 minutes after it fell.
The snow in the area was only a foot deep, but the wolves were sinking in about 6 inches.
8. (February 7, 1960. About half a mile southeast of the south west end of Angleworm Lake.) At 4:10 p.m. the pack of 16 headed north from Moskey Basin for a quarter-mile on an old wolf trail. At 5 o'clock the animals suddenly veered upwind and became alert, often stopping and pointing or scenting the wind. All wolves stayed close together and did not dally. They traveled three-quarters of a mile to within 250 yards of a cow and calf which were browsing directly upwind (5:30 p.m.).
The wolves gave no indication of scenting the moose. Instead, they turned through a thick spruce swamp; but when a third of the way across, they suddenly headed toward the moose. As the pack approached to within 100 yards, the moose started running, the cow behind the calf. The wolves gave chase and soon were racing alongside and behind them.
Throughout the chase, the cow defended the calf, charging the wolves frequently. One animal managed to bite the calf's rump once but did not hinder the animal. The pursuit continued for 200 to 300 yards (through many types of cover and over varying terrain) without an attack, but eventually the wolves separated the moose. Most of the pack pursued the calf, while two animals followed the cow. After a chase of several hundred yards more, a few wolves attacked the rump and flanks of the calf; one grabbed it by the left hind leg. The cow caught up with the group and managed to stamp on one wolf, which arose instantly and appeared unhurt. The others released the calf and continued pursuing it for another hundred yards before attacking again. They finally pulled the animal down and tore at it, but it arose and the cow rushed in. Some of the wolves fled, but others chased the cow. Then the wolves assailed the calf once more. One grabbed it by the nose, and three or four tore at its neck and throat; others ripped at its rump. The calf's hind quarters went down, but the animal continued on, dragging its hind legs and the wolves that were attached to its body. It managed to stand once more, and the cow started to charge again, but one wolf chased her away.
The wolves made a final attack on the calf, and it was unable to arise. Then they lined up side-by-side around the carcass and began feeding. The cow gradually wandered back toward where she had been jumped.
13. (February 12, 1960. About 1-1/2 miles southwest of Halloran Lake and about 200 yards northwest of the Isle Royale shore.) The 16 wolves were traveling along the shore when suddenly they veered inland about 2:30 p.m. toward a lone cow (sex determined later) standing on a ridge 200 yards upwind. The animal ran when the pack was 100 yards away, and the wolves charged up the ridge and continued on her trail. The cow ran slowly and stopped to look back at the approaching pack, which caught up within 100 yards. She stood next to a bushy spruce for protection, and as the wolves lunged, she charged and kicked at them with all four feet. Although she seemed to connect with her hind feet, apparently no animals were injured.
Meanwhile, the whole pack caught up. The moose defended herself for about 3 minutes while backed against the spruce, but suddenly she bolted and fled toward the end of the ridge. The wolves attacked her rump and flanks but released their holds as she brushed through some thick spruces. They pursued the animal for 25 yards to the end of the ridge, where all plunged down the steep slope.
When the moose landed at the base of the ridge, the wolves were attached to her back and flanks, and one held her by the nose. The downed animal attempted to rise, but the sheer weight of the wolves seemed to anchor her. The wolf grasping her nose held on firmly while she violently shook her head. Most of the animals continued working on her rump and flanks, while two tore at her shoulders.
The moose struggled for more than 5 minutes while the wolves, packed solidly around her, tugged away. Two individuals had to wait at one side, for there was no room around the moose. The "nose-wolf" continued its hold for at least 10 minutes, while the others pulled from all sides. After about 10 minutes, the moose appeared dead. This cow was in wear-class VI.
14. (February 15, 1960. About half a mile downstream of the junction of the Grace Creek Trail and Grace Creek.) At 2:10 p.m. the 16 wolves were heading down Grace Creek. Suddenly, they pointed toward a cow (as determined later) 200 yards to their left. Then they continued down the creek to where it wound closer to the moose. Heading inland over a knoll, the wolves surprised the cow 25 yards away.
The animal fled, but the wolves caught up almost immediately. One grabbed her right hind leg just above the hoof. However, as the cow trotted through some spruces, she shook the wolf loose. She then ran in a semi-circle toward the creek (figure 91), and several times the wolves overtook her but failed to attack. Once when she ran through a snowdrift, the wolves lost ground, but they quickly caught up again.
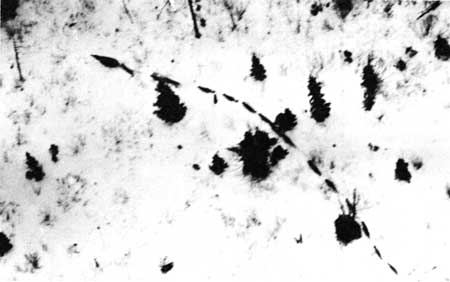
Figure 91—Wolves pursuing a moose near Grace
Creek.
As the moose started down a shallow valley, the wolves attacked her rump. She soon shook them, however, and proceeded to the frozen creek bed, where the wolves attacked again. One animal kept jumping at her nose and finally grabbed it; others fastened onto her rump and flanks. The cow fought hard and dragged the wolves about 100 yards downstream (figure 92). Three or four times, she lifted the "nose-wolf" off the ground and swung it for several seconds before lowering her head. This wolf maintained its grip for over a minute. The moose continued fighting hard and finally shook the wolves and ran back upstream, with the whole pack following.

Figure 92—Wolves attacking a moose. Note that
one wolf has the moose by the nose (arrow).
The cow started into the woods and the wolves lunged again. The moose kicked constantly and trampled two individuals into the snow. One of them crawled away but later seemed unhurt. The moose then stood next to a small balsam along the creek shore and continued to fight off the wolves, which soon gave up temporarily and lay on the ice. At 2:35 p.m. they went 200 yards downstream and assembled. They returned to the animal three times but found her belligerent, although blood from her wounded rump covered several square yards of snow. Nevertheless, there appeared to be no mortal wound.
From 2:50 to 3:25 p.m. the wolves lay on the nearby ice. Meanwhile, at 3:20 the moose walked about 10 yards and lay down. At 3:25, the pack approached and she arose again. Although appearing stiff, she charged the wolves effectively. Many of them were eating the bloody snow where she had stood first (figure 93). At 3:40 the wolves lay down again and at 3:50 so did the moose. About a minute later, a wolf approached the moose and she arose again. At 4:12 this occurred once more. Then the wolves entered some spruces 25 yards south of the moose and curled up. From 4:20 to 4:40 we were refueling, but when we returned, the wolves were still there.

Figure 93—The large pack waiting for the
wounded moose to weaken. See Hunting Account 14. (c) National
Geographic Society, courtesy National Geographic Magazine.
At 5 o'clock, they arose, tested the moose, and found her quite pugnacious. Ten minutes later, 14 of the animals left and headed southward while 2 remained curled up within 25 yards of the wounded moose, which was also lying down.
From 5:35 to 6:05, the pack visited an old kill half a mile south of the creek; the animals then traveled back along a ridge until half a mile from the wounded cow. Meanwhile, the two "guards" arose and stood near the moose. The pack headed almost directly toward them, and at 6:40 p.m. when we had to leave, the pack was within a quarter-mile and still heading toward the wounded moose. The next morning at 10:50 a.m. the wolves were feeding on the carcass, which was where we had seen the live animal. Ground observation later showed that this cow was in wear-class VI.
40. (March 17, 1960. About 50 yards southwest of the Island Mine Trail and about half a mile from Siskiwit Bay.) At 11:10 a.m. the large wolf pack was resting along the trail, and at 11:25 the animals slowly headed 150 yards farther up the trail. At 11:35 they suddenly turned upwind and ran about 50 yards into a thick spruce stand. Two moose ran through the stand and split up. Because of thick cover, we could not see the wolves.
Suddenly, however, a calf, pursued closely by two wolves, headed out of the stand, down the trail 100 yards, and into a spruce swamp on the other side of the trail. Within 100 yards, the wolves began nipping at the hind legs of the moose. After another 50 yards, one wolf was clinging to the animal's rump and the other to its throat. The moose stopped and trampled the front wolf, but the wolf would not let go. It clung to the calf's throat for about 2 minutes while the calf continued to pound it and drag it about.
Finally this wolf released its throat-hold, but the other still stuck to the rump. The first wolf then stood on its hind legs, and placing its front paws on the left side of the moose, started chewing the side of its neck for several seconds. The calf soon brushed this animal against a tree, but the wolf then dived under the moose and fastened to its throat. As the running moose straddled the wolf, the wolf ran along with it for about a minute.
Meanwhile, two other wolves caught up. One bit the calf around the head and finally grasped its nose. The other grabbed the right flank and then changed to the rump where it clug for about a minute while the moose continued on. This, one wolf had the calf by the nose, one by the throat, and two by the rump. The animal soon stopped and was pulled down under a small clump of trees. In about 3 minutes, it ceased struggling (11:45 a.m.).
When the two wolves first attacked the calf, the cow tried to catch up with it but was too far behind the swamp to find it. Total distance of the chase was about one quarter of a mile. Apparently, the rest of the animals had been chasing the cow; eventually they found their way to the calf also and joined in the feed.
41. (August 26, 1960. West shore of the north arm of Chickenbone Lake about 75 yards south of the outlet.) At 6 p.m. on August 23, four campers noticed a cow moose about 30 feet inland of the above location with a large open wound on her left hind flank. She seemed reluctant to move. Two days later, Chief Ranger B. J. Zerbey reported that the moose appeared sore and short of breath and would not arise. Two nearby bloody beds indicated that she had arisen a few times but had been hesitant, or unable, to leave the area. At 6 p.m. the same day, two fishery biologists saw her in the mud at the edge of the lake and thought she acted lively. However, the next morning at 8 o'clock, as these men approached the area, they heard wolves barking. Rowing by in their boat, they saw that the moose was dead.
At noon I examined the carcass. There was a surface wound about half an inch wide on the left cheek, and several long gashes on the throat, but none of these had bled much. Horizontally across the upper left hind leg was a wound about 2-1/2 inches deep, 4 inches wide, and 8 inches long. The exposed muscle hung ragged, appearing well chewed; undoubtedly, this was the wound observed by the campers, Zerbey, and the biologists. The only other exposed area was the pelvic region. The flesh there had been eaten through to the coelom, and a few loops of intestine were pulled from the body. Probably most of this damage resulted from feeding, for none of the observers mentioned wounds in this area although they did notice the less conspicuous upper-leg injury.
This cow was in wear-class V and harbored 57 hydatid cysts in her lungs.
During the next 2 days, I saw (from an aircraft) at least three, and possibly four, wolves on a nearby ridge, and heard at least four. These may have represented both small packs or part of the big pack.
71. (March 12, 1961. About 1-1/2 miles southeast of the northeast end of Lake Desor.) From March 6 to 11, the aircraft underwent its 100-hour check, so no aerial work was done until the 12th. About 10 a.m. the large pack was found at a fresh kill southwest of Halloran Lake. Tracks showed that the last kill had been near Fisherman's Home. While backtracking the wolves from there, we saw a lone wolf also backtracking them along the northwest shore of Siskiwit Bay at 10:30 a.m. We continued following the trail to the above location, where a badly wounded moose lay on an open hillside. Tracks showed that the large pack had wounded this animal, stayed around for at least several hours and then abandoned it. This probably happened about March 8.
At 12:20 p.m. we saw the lone wolf 1-1/2 miles from the moose, still backtracking the pack. From the animal's attitude when approaching I had little doubt that it knew the wounded moose was there. When 250 yards away the wolf ran excitedly up the trail but became cautious when 50 yards away and circled to the west to approach. I learned later that the moose was a bull. He was lying in a blood-soaked patch of snow about 15 feet in diameter which he had not left since the attack. As the wolf came within 30 yards, he arose (12:55 p.m.). The wolf approached to within 10 feet, circled the bull a few minutes, and went off 30 feet and lay down (figure 94). After 5 minutes, the moose lay down; immediately the wolf ran to him, so the moose stood (figure 95). The wolf lay down about 20 feet away, and 10 minutes later the bull lay down. Again the wolf threatened him, tail wagging excitedly, and seemed to try for his nose but failed. The moose just stood without moving quickly or threatening the wolf. A few minutes later the wolf lay down again. The bull continued standing at least from 1:20 until 1:45 p.m. when we left to refuel.

Figure 94——A lone wolf waiting for a
wounded moose to weaken.

Figure 95—The wounded moose stands upon approach
of the wolf.
From 2:45 to 3:30 the wolf lay sprawled on its side about 20 feet from the moose, which continued standing.
At 4:45 the bull was alive but lying down, and the wolf was tugging at his rump. Intermittently the moose watched the wolf but made no threats. He seemed to have no feeling in the rump, or more probably, he was too weak to stop the wolf. At 5:55 the moose was still alive, but by 6:30 p.m. he was dead, lying on his right side.
The next morning at 11 o'clock, I examined the carcass. The only apparent wounded areas were the rump and thighs, but the pelvic region had been fed upon, so it also may have been wounded. The animal was a bull in wear-class VIII and was heavily parasitized with ticks and hydatid cysts, although the femur marrow was normal.
DISCUSSION OF KILLING TECHNIQUES
In all kills witnessed, the first point of attack was the rump. In fact, this region was the only site of severe wounds on the two adults examined from the ground (figure 96). During 1956 and 1957, Cole (1957) observed adult moose on Isle Royale wounded in the thighs. In Alaska, Burkholder (1959:9) chased off a pack of wolves attacking a moose. The animal died that night and was examined from the ground the next day. "The only injuries observed consisted of deep bites and tears on the hams above the hocks. The animal was not hamstrung, the tendons being still intact. No other wounds were noted." Cowan (1947:159) examined several elk kills and in each instance the attack had been from the rear and side with the wolf seizing the flank at the point where the leg joins the abdomen." In Minnesota, a moose was reported wounded on the hind flanks by wolves (Stenlund, 1955).
Figure 96—Freshly killed moose showing wounded
area.
Young and Goldman (1944) reported that wolves hamstring their prey and eat out the hams. However, no evidence of hamstringing was found during the present study; and Burkholder, Cowan, and Stenlund each stated specifically that they observed no instance in which wolves hamstrung their prey. During caribou studies in the Northwest Territories, Banfield (1954:47) found that hamstringing was seldom done. "The method that the wolf generally uses for killing a caribou is to race alongside of it and pull it down by grasping the flank, shoulder, or throat, with the jaws." Stenlund (1955:31) gave the following description of killing techniques used on deer:
No evidence of hamstringing of deer was found on freshly killed carcasses although the possibility does exist. Usually deer are run down from behind, the wolf or wolves biting at the hind flanks and abdomen, or at the hind flanks and head region simultaneously. Often the deer is knocked to the ground two or three times before it is killed. It is possible that some deer might even die from a combination of shock, fright, and exhaustion rather from actual wounds since in some cases it did not appear that animals were wounded badly enough to cause death.
Five of the six times I watched wolves wound moose, several animals slowed down the prey and occupied its attention by pulling at its rump, and then one wolf grabbed the nose (figure 92). During the sixth hunt, trees obscured my view after the wolves attacked the animal's rump. Although the nose hold is not mortal, it stops the prey and distracts it from the wolves on the rump. In some hunting accounts furnished by Young and Goldman, mention is made of part of the pack distracting the prey at the front while others inflict significant damage to the rump.
Although the rump region contains no especially vital parts, it seems to be the least dangerous and most advantageous attack point on such a large animal as a moose. After the wolves rip through the 5-inch hair and thick hide, every injury to the upper-leg musculature would hinder the movement of the prey and render the animal more susceptible to intensified attack. Once downed and besieged by several wolves, the moose succumbs quickly. Each of three calves and one adult was killed within 10 minutes.
Apparently many adults are wounded, left to stiffen and weaken from their rump wounds, and then killed (figure 93). Three instances were observed in which this tactic was employed, and Cole (1957) reported two. Probably bulls attacked by the large pack, and any adults tackled by the smaller packs, are likely to be victims of this tactic. This probably is not done purposely; rather it appears to result from the wolves' failure to wound a moose sufficiently to incapacitate it.
Sometimes when the wolves wound a moose, they cannot complete the kill, so they abandon the animal. The moose probably dies within a week, and there are indications that the wolves return and feed on the carcass. The animal involved in Hunting Account 2 was abandoned on February 24, 1959, but on March 13 the pack was feeding on a kill within a quarter-mile of where the animal had been left. Of course, there is no certainty that this moose was the one abandoned. An observation on March 15, 1960, also may be significant. After we followed the large pack all day, the animals began exploring a swamp southwest of Halloran Lake. They did not appear to be hunting but seemed to be searching for something. Eventually they discovered a calf carcass beneath a clump of cedars and began feeding. The femur marrow of the animal was fat-depleted and the stomach was full of cedar. Since this area is heavily hunted by the wolves, it seems unlikely that a moose would get a chance to starve to death. It is more likely that the wolves had wounded and abandoned the animal in this area, and that the only available browse was the small amount of cedar from surrounding trees.
On another occasion, when the wolves had not eaten for 4 days, we followed them all day until they excitedly entered a small clump of trees. Although there was no chase, the animals raced to the clump and gathered there. After about 45 minutes, a few left the clump and lay in the open for the rest of the day.
The next day we found that they had fed on a carcass under the trees. Either they killed the moose while we circled above (doubtful, since their actions did not indicate this), or they found the carcass. The femur marrow was normal, so the animal was not in a state of extreme emaciation. Perhaps it had been wounded and abandoned weeks before.
The moose involved in Hunting Account 72 undoubtedly would have died within a few days if the lone wolf had not finished it off, for it remained for days in a small area, which soon was completely browsed out.
MOOSE DEFENSE
Regarding the senses of moose, Peterson (1955:102) found that ". . . the ears often serve to alert the animal, the eyes to investigate, while the final stimulus, causing immediate reaction, is transmitted by smell." Moose behavior during the present study indicated the same. Since wolves generally traveled upwind to their intended prey, they often approached to within 100 yards before discovery by the moose. Under certain conditions, moose sensed wolves when a quarter of a mile away, but on one occasion the pack came within about 5 feet of two moose in their beds. Differences in wind direction and velocity, cover, terrain, and other factors probably accounted for the varying abilities of moose to sense wolves.
No observations were made on the summer defense of moose against wolves, but two reports indicated that the animals seek refuge in water when threatened. In 1958, campers related to D. L. Allen that they saw a cow and calf enter Tobin Harbor at its head. Then three wolves emerged from the woods, paced the shore about three times, and returned into the brush. A similar occurrence was noticed by Mrs. Alfreda Gale of Tobin Harbor and H. T. Orsborn of Rock Harbor. Mrs. Gale reported that, in July 1959, she and Orsborn spotted a moose "prancing and snorting" about 20 feet from shore in Gutt Bay, Tobin Harbor. Then they spied a wolf on shore near the moose and were convinced that the moose was taking refuge from it. After a few seconds, the wolf ran off, but the moose remained in the water for several minutes.
Sign around remains of a calf killed by wolves about August 5, 1960, indicated that the moose had been killed in the shallow water of an old beaver pond. Apparently, it sought protection in the pond but did not reach deep water. Cowan (1947:160) reported that "on several occasions single deer, elk, and moose have been seen making use of this defensive behavior . . .," and Peterson (1955:104) agreed that ". . . moose regularly make for the nearest water when seeking protection from predators."
In seven hunts (11 animals) witnessed in winter, moose detected wolves before being discovered, and in each case, the moose immediately left the area. Since it will be shown that a low percentage of moose tested by wolves are killed, it is safe to assume that most of the above-mentioned moose were not highly vulnerable and that, nevertheless, they chose to avoid an encounter with the predators.
During 36 hunts involving 73 moose, the animals fled without being attacked (table 16). They all were pursued, but either obtained adequate headstarts, outdistanced the wolves, or outlasted them. Soon after the wolves discontinued a chase, the pursued animal stopped and watched its backtrail. Several times moose in such situations appeared to be saving their strength until hard pressed. Murie (1944) noticed that caribou frequently stood around watching nearby wolves ". . . when they could have been moving away to a more secure position." Perhaps this was for the same reason, for caribou also depend upon speed and endurance to escape wolves.
TABLE 16.—DEFENSIVE BEHAVIOR OF MOOSE ENCOUNTERED BY LARGE PACK OF WOLVES OR PART OF ITa
[Parentheses indicate the number of hunts involved]
| Year | Number of moose employing each defense | ||
| Ranb | Ran, then stood |
Stoodc | |
| 1959 1960 1961 |
12 (4) 27 (12) 34 (20) |
0 8 (7) 4 (4) |
2 (1) 11 (7) 11 (9) |
| Total | 73 (36) | 12 (11) | 24 (17) |
bProbably more in this category would have stood, if wolves had been close enough.
c More of these animals might have run, if wolves had not been so close.
Even when the wolves overtook their quarry, in many cases the moose continued running until their pursuers tired. In seven instances (nine animals) moose ran at least half a mile before the wolves abandoned chase. Three animals, including a cow and calf, traveled about 2-1/2 miles, and another cow and calf ran approximately 3 miles before the wolves gave up. During long pursuits, even after the pack stops, the moose usually continue running for at least a quarter of a mile.
The gait of a moose chased by wolves appears effortless; the animal takes long, deliberate, trotter-like strides. The longest distance I saw moose maintain this gait was an estimated 3-1/4 miles. This was accomplished by a cow and calf closely pursued by the large pack on March 11, 1960, and it helped them escape. Although healthy moose apparently can outlast wolves, they cannot out-man them. Once wolves catch up to a moose, they usually remain even with the animal unless deep snow or tangled blowdown interferes. However, the maximum speeds of the two species appear to be similar, for if a moose maintains about a 100-yard lead for 15 or 20 seconds, the wolves soon give up. Reported speeds for the moose are: 19 m.p.h. (Peterson, 1955), 22 m.p.h. (Findley, 1951), 27 m.p.h. (Cowan, 1947), and 35 m.p.h. (Cottam and Williams, 1943); a moose maintained the latter speed for a quarter of a mile. Peterson reported that moose gallop occasionally when frightened, but I did not observe this gait used by any moose pursued by wolves.
Running moose were not hindered by blowdown, thick swamps, or 3 feet of snow. Peterson (1955) found that 30 inches of encrusted snow presented little hindrance to moose, but cited findings by Wright et al. that crusted snow lacerates their legs, seriously hampering travel. Murie (1944) also mentioned this hazard. Since strong crusts apparently did not form on Isle Royale until early spring, no observations were made on this subject. However, in late March and early April, crusty snow might be a significant factor in wolf-moose relationships. In British Columbia, changing snow conditions were found to affect predation on moose profoundly. Stanwell-Fletcher (1942) reported that wolves did not hunt moose "in earnest" until the end of January, when 6 to 8 feet of snow impeded travel by the latter.
Since so many moose escaped by running (even those which the wolves overtook and chased for more than a mile), running appears to be a successful defense. However, each of the five mortal attacks witnessed from the beginning involved running animals. Perhaps this is because a moose does not employ the most effective weapons, its hoofs, so easily when running as when standing still, and because weak or inferior animals, which may be afraid to defy wolves, probably are among those moose that flee. Since any weakness probably affects the ability of a moose both to run and utilize its hoofs effectively, the wolves soon might sense the animal's debility.
Twelve moose, involved in 11 hunts, fled as wolves approached but stopped before, or as soon as, the pack caught up. The moose then stood at bay and held off the wolves. In 17 other instances, 24 moose stood their ground immediately, including 4 cows with calves. In addition, a cow and calf stood at first and then defensively strode off with wolves following for about a quarter of a mile.
None of the 24 moose which stood at bay throughout an attack was killed or wounded. I do not know whether this is because only strong, healthy, "confident" animals defy their attackers or because defiant moose are so formidable. Probably the reasons are of equal importance. It is obvious that moose which stand their ground when confronted by wolves enjoy full command of the situation (figure 1). On February 28, 1961, I saw a moose stride boldly for about 70 yards to meet seven approaching wolves which turned and left when the moose was about 30 yards away.
It is easy to see why wolves fear an enraged moose. With mane erect, ears flattened back, neck extended and head held low, the surrounded animal lashes out at the nearest wolf, and wheels quickly to chase any individual which dares close in behind. Both fore and hind hoofs are used with great facility and exactness. Sometimes a harassed moose backs against a conifer or other protection for its rear. Because my observations were from an aircraft, only the conspicuous manifestations of rage were seen, but Denniston (1956:111) provided a more complete description of the rage pattern:
In this pattern the mane is erected, the ears flattened back against the neck, the lips retracted, the tongue protruded and curled up over the upper lip and nose and repeatedly darted in and out licking the upper lip. The animal usually rears on its hind legs pawing the air with the forefeet, if the stimulus object is at a distance of less than about forty feet.
An informative close-up account of defensive behavior of an Isle Royale moose against a dog is furnished by Hickie (n.d.:28):
The cow . . . charged Togo, ears back, head close to forelegs with outstretched neck and bristling mane, and both fore and hind feet stamping at every jump—and how she came! We barely had time to get around behind a cabin before she charged past. Togo tried to duck to the side, but the heavy snow was too much for him and he floundered. I thought it was all up with him, for she caught him and pounded him with her front feet; but as luck would have it, her aim was poor and he ran toward me. She wheeled and caught him, again rolling him in the snow. This time he managed to roll back under her front legs, scramble out to one side and run down towards the house. After this Togo stayed his distance and as soon she laid back her her ears and moved towards him, he was gone like a flash down the trail.
I once watched a moose beat two wolves into the snow, but these escaped unharmed; apparently the blows were softened by the snow. In another instance a 9-month-old calf pounded a wolf clinging to its throat and finally persuaded the animal to seek a less-hazardous hold; but the wolf was not hurt. In British Columbia, a large male wolf was found barely alive, with broken ribs and legs. "Surrounded by moose tracks, blood patches and moose hair, the wolf had been cripped in a great battle" (Stanwell-Fletcher, 1942:138). In the Canadian Northwest Territories, MacFarlane (1905) found a live adult wolf with a hind leg shattered by a kick from a bull moose.
When a moose stands defiantly at bay, the wolves try to force it to run, but unless it does, they abandon it. Once when a moose stood its ground, the wolves left within 30 seconds, and the longest the pack harrassed an uninjured standing moose was 5 minutes. Cowan (1947) reported instances of a cow elk standing off seven wolves and of a moose discouraging three wolves by standing and striking with its forefeet. Additional evidence that Isle Royale wolves fear moose at bay was afforded by five observed hunts in which the pack chased one of a group of moose until it stopped, and then immediately pursued one of the other fleeing animals. Even when a moose is wounded but throws the wolves and stands its ground, the wolves fail to continue the attack. In each of four instances observed in which a moose was only wounded, the animal finally stood at bay. The wolves then waited for it to weaken considerably before attacking again. In two of the cases they eventually left the wounded moose. This evidence emphasizes that when a moose stands its ground, the wolves are reluctant to attack.
Defense of the calf in winter is quite strong and sterotyped. If wolves are detected soon enough, the cow leads her calf quickly away. If the wolves pursue in such a situation, or if they surprise the moose, the cow immediately rushes to the rear of the calf. In four observations, the two moose stood at bay, and the wolves left within a minute. While standing off a pack of wolves, the cow makes short charges at the animals and instantly returns to the calf's rump. If necessary, the calf employs its front feet for defense, but these probably do not afford adequate protection. In each of the observed cases, the cow provided the effective defense.
In eight hunts the cow and calf ran from the wolves, and in six of these the wolves pursued closely for long distances, once for about 3 miles. During such chases, the moose run slowly and deliberately, with the cow staying close to the calf's rump. If any wolf threatens the calf, the cow charges it but immediately returns to the calf. When the two animals are together they constitute an invulnerable team. The calf charges wolves in front of it, and the cow protects the rear and flanks of the calf and kicks any wolves at her rear. However, while traveling through varying cover and over widely dissimilar terrain, the two cannot always remain together. If a calf gets a few yards from the cow, the wolves close in; if the cow does not rush in promptly, the calf is doomed. Each of the three calves which I saw killed was separated from its parent while running.
Little is known about the summer defense of the calf. Murie (1944) described a cow standing and protecting her newborn calf from two wolf-size huskies. Peterson (1955) discussed the close relationship between cow and calf, and related personal observations of the strong protective instinct of the cow. (For a discussion and analysis of the strong cow-calf bond, see Altmann, 1958.) During the first week or two of a calf's life, the cow would have to stand off wolves instead of leading the calf away, for the calf could not run fast or far enough. On May 22, 1961, I startled a cow and very young calf about 100 feet away. The calf failed to keep up with the cow, so the cow had to wait for it. At times, the calf dropped about 25 feet behind. If the cow were to behave in this manner with a wolf in pursuit, the wolf probably could catch it before the cow could interfere. Undoubtedly, a single bite from a wolf would incapacitate a young calf. Had I been a wolf, however, the cow probably would have stood her ground, or at least remained close to her calf.
An encounter on June 13, 1961, demonstrated the attitude probably assumed by a cow and calves retreating from wolves. While I stood on a moose trail photographing moose in Ojibway Lake, a cow and twin calves ambled down the trail behind me, about 30 feet away. Since there were no nearby trees to climb, I motioned to alert the animals to my presence before they got too close. The cow stopped, grunted, and laid back her ears, and the calves huddled around her hind legs. She acted quite "confident" while I fumbled with my camera. Then she turned and headed slowly and deliberately away, the calves remaining at her heels.
W. Leslie Robinette reported an incident to me which showed what might be in store for wolves which attempt to bother a calf. While studying moose browse in the park, on May 19, 1961, Robinette happened upon a cow and newborn calf (with umbilicus still apparent) south of Siskiwit Lake. After observing the pair from one tree for 45 minutes, he climbed another for better visibility. Each time he changed position, the cow snorted and her mane bristled. When he descended the second time, the cow charged. Robinette ducked behind the nearest tree just as the animal straddled the tree and pounded the ground on each side of him; her nose was but a few inches from his. Immediately after the initial attack, the cow returned to her calf.
PREDATION EFFICIENCY
Many biologists have suspected that predators have a low rate of hunting success, but quantitative evidence for this belief is not easily obtained. One of the few studies comparing figures on successful and unsuccessful hunts by any predator is Rudebeck's (1950, 1951). By recording observations of all hunts by four species of European raptores for 5 years, Rudebeck found that only 7.6 percent of 688 attempts to secure prey were successful. Success percentages for individual species varied from 4.5 percent to 10.8 percent.
The present study provided the opportunity to obtain figures on the rate of success, or "predation efficiency," of the large pack of wolves (table 17). A total of 160 moose were estimated to be within range of the hunting wolves while under observation, but only 131 were detected. Of these, 77 were tested by the wolves; i.e., the wolves chased them or held them at bay, so those which escaped did so because of their superior condition or ability. Those which were detected but not tested also may have escaped on this basis, but circumstance probably was more important. Therefore, predation efficiency is considered here as the percentage of animals tested that are killed. Since 6 moose were dispatched out of 77 tested, the predation efficiency is 7.8 percent. The almost-exact agreement with Rudebeck's figure undoubtedly is coincidental, but the fact that both percentages are of the same order of magnitude is notable.
TABLE 17.—RESULTS OF HUNTS BY THE LARGE PACKa
[Figures in parentheses indicate number of hunts involved]
| Year | Total hunts |
Number of moose | ||||
| In range | Detected | Tested | Wounded | Killed | ||
| 1959 1960 1961 Total |
7 44 34 85 |
16 83 61 160 |
15 (6) 66 (33) 50 (27) 131 (66) |
7 (5) 48 (26) 22 (15) 77 (46) |
2 (2) 5 (5) ... 7 (7) |
1 (1) 5 (5) ... 6 (6) |
| Percent in range Percent detected Percent tested |
81.9 .. .. |
48.1 58.8 .. |
4.4 5.3 9.1 |
3.7 4.4 b7.8 | ||
b"Predation efficiency."
PATTERNS OF SELECTION IN THE MOOSE KILL
Several other authors have reported that wolves make many vain attempts to secure prey (Murie, 1944; Cowan, 1947; Harper, Ruttan, and Benson, 1955; Crisler, 1956), although Burkholder (1959:9) stated that "there was no evidence during this period of my study to indicate that wolves, even singly, had a difficult time catching adult caribou." After extensive study of wolf-Dall sheep relations, Murie (1944:109) concluded that
Many bands seem to be chased, given a trial, and if no advantage is gained or weak animals discovered, the wolves travel on to chase other bands until an advantage can be seized. The sheep may be vulnerable because of their poor physical condition, due to old age, disease, or winter hardships. Sheep in their first year also seem to be specially susceptible to the rigors of winter. The animals may be vulnerable because of the situation in which they are surprised. . . . My general observations indicate that weak animals are the ones most likely to be found in such situations.
Murie also witnessed several caribou hunts in which wolves appeared to be testing a herd for weak animals. Regarding predation on moose, Murie (1944:186) stated
Wolves perhaps worry many moose which fight them off with such vigor that they are unwilling to expose themselves to the deadly hoofs. However, if any sign of faltering is shown, due to old age, food shortage, or disease, the wolves would no doubt become aware of it, and one would expect them to become more persistent in their attack in hope of wearing down the animal. Moose which are actually known to have been killed by wolves should be closely examined to determine their condition. Unfortunately in many cases the evidence is destroyed.
Since such a low percentage of moose tested are killed, wolves probably are selecting certain types of individuals. These could be weak or inferior moose, or merely animals in unfavorable situations. The latter possibility seems unlikely, for observations have been made of unsuccessful hunts occurring on several types of terrain, in various cover, and in diverse situations.
Information on sex, age, and condition of wolf-killed moose was obtained from remains found both from the aircraft in winter and from ground search in summer, as described on page 115.
Sex and Age Distribution. When possible, sex and age data were secured from all moose remains. Sex determination was based on the presence or absence of antlers or antler pedicels. The sex ratio of winter wolf kills was 22 females to 11 males, indicating a strong selection for cows, at least during February and March (assuming an even sex ratio in the population). However, the sexable remains found by ground search in spring and summer, which should include year-round mortality, showed a ratio of 18 females to 27 males. If there has been an even sex ratio in the population for the last few years, the number of males and females in this category of remains should be even. The preponderance of males might be caused by the probability that the more massive male skulls remained intact longer than female skulls; thus, more females than males would be classified "unknown." Since many of the remains found by ground search consisted only of old, bleached bones, a high proportion of the sample includes animals dead several years. If data from animals which probably died before 1955 are eliminated, the sex ratio is 8 females to 7 males.
Age estimates were based upon mandibles, toothrows, or molariform teeth collected from the remains. Specimens of like molar-wear patterns were segregated, and 10 classes corresponding to those described by Passmore et al. (1955) resulted. These authors estimated that classes I to IV correspond to the specimen's actual age but that classes V to IXA contain specimens varying 2, 3, or more years in age; e.g., class VII might contain specimens from 8-1/2 to 10-1/2 years old. However, Sergeant and Pimlott (1959) aged moose on the basis of annuli found in sectioned incisors, apparently a more precise and accurate method. They compared age estimates provided by Passmore et al. with their own for the same wear classes, and concluded that both variation in age, and maximum age, were greater in most classes than formerly thought; e.g., class VII included specimens 10 to 17 years old. In the present study, the ages furnished by Sergeant and Pimlott for each wear class were accepted as more representative of actual ages of the specimens.
The similarity in age composition between remains representing year-round mortality and those of winter wolf kills can be seen in table 18. The greatest bias undoubtedly occurs in the calf class of the year-round sample, for calf remains would be hardest to find and would disintegrate sooner. Nevertheless, both samples indicate that calves bear more losses than any other class. This is expected merely because calves form the largest single class. To demonstrate that wolves select calves, one must prove they kill a larger proportion of calves than exists in the population. Since calves composed a 3-year average of 15 percent of the sampled winter population but 36 percent of the 3-year sample of winter kills, a definite selection for calves is indicated, at least in that season.
TABLE 18.—AGE DISTRIBUTION OF DEAD MOOSE
| Wear classesa | Estimated ageb (years) |
Number of winter wolf killsc |
Number of remains found by ground searchd |
| Calf | .. | 18 | 18 |
| I | 1 | .. | 1 |
| II | 2—3 | .. | 2 |
| III | 3—4 | .. | 1 |
| IV | 4—7 | .. | 4 |
| V | 6—10 | 3 | 8 |
| VI | 8—15 | 15 | 8 |
| VII | 10—17 | 3 | 6 |
| VIII | .. | 5 | 5 |
| IX | .. | .. | 4 |
| IXA | e 20? |
6 | .. |
| Total | .. | 50 | 57 |
bSergeant and Pimlott (1959).
c Found from aircraft. See page 115 for qualification.
dIn spring and summer.
e Passmore et al. (1955:238).
In Alaska, Burkholder (1959) found that six of seven ageable, winter wolf kills were calves, and one was a yearling.
During summer, calves seem to compose even a higher proportion of the kill, if scat-analysis figures are valid indicators (table 10). In scats from 3 summers, there were 162 occurrences of moose remains identifiable as calf or adult; of these, 75 percent were calf remains. (These statistics might be biased toward calf remains because calves have a higher proportion of hair—the primary identifiable remain in scats—than do adults. Also see page 164.) In addition, three of the four moose found fed upon by wolves in summer were calves. Cowan (1947:167) reported that, in British Columbia, all remains of moose in the summer wolf scats he examined were calf remains. However, he cautioned that moose calves are more prone to accidents than are the young of other big game, and that "carrion may well make up a fair part of the calf moose item."
Calves probably are especially vulnerable to wolf predation because, like other growing animals, they are smaller, weaker, less experienced, and less independent than adults. In winter, calves may be heavily infested with ticks (see page 107), which probably would predispose them to predation. Fenstermacher (1937) reported that ticks are particularly debilitating, and Peterson (1955:186) believes that ". . . the most serious effects of tick parasitism are manifested in the reduction of the vitality of the moose, making them more vulnerable to other factors, such as diseases, predation, abortion, and malnutrition." In British Columbia, Hatter (1950a) discovered that calves constituted two-thirds of 161 moose found dead from tick-malnutrition complex early in 1947.
Since calves depend so completely on their mothers for protection from wolves, the condition of cows also is all-important to calf survival.
The most significant information in table 18 is that most moose in wear-classes I to IV are invulnerable to wolf predation. This is obvious in the winter wolf-kill sample, but the year-round category also strongly indicates the same. The few specimens in classes I to III of the latter sample could be results of accidents, for animals of any age are susceptible to accidental deaths.
It is surprising that yearlings are so secure from wolf predation. In February and March, short-yearlings are killed despite their mother's protection, but apparently by June, when they become independent, they seldom are taken. Many yearlings in June seem no larger than they do in March, so size probably does not make the difference in early summer. Possibly the primary reason that few yearlings are killed is that wolves change their hunting habits. Just after yearlings are forced away by their parents, a new crop of calves is produced, and wolves undoubtedly prey on these throughout the summer. Meanwhile, the yearlings grow quickly and by August probably are large enough to protect themselves. Murie (1944) found that wolves in Mount McKinley National Park preyed heavily upon caribou calves and took only a few adults during the summer.
Moose in wear-class VI, estimated at 8 to 15 years old, seem to be preyed upon most. Of course, individuals older than these would be as vulnerable, or probably more so, but since most kills are in the 8- to 15-year-old class, few older animals remain. Even if a moose population were not hunted by man or wolves, one would expect most adult mortality to occur in the oldest age classes.
No figures are available on the age distribution of moose mortality in other areas for comparison with the Isle Royale statistics. However, Peterson (1955:176), without furnishing figures, stated that "a great majority of the skeletal remains of moose found in the St. Ignace area were of old animals with well-worn teeth." Sources of mortality included wolf predation.
In Mount McKinley National Park, Murie (1944) established that over 50 percent of adult Dall sheep mortality occurs in ages 9 to 14 years. Tener (1954) found that, in the Canadian Northwest Territories, 21 of 24 adult musk-ox remains were from animals at least 6 years old. He believes that many of these had been killed by wolves. During a study of wolves and deer in Minnesota, Stenlund (1955:43) concluded that "there is no indication that wolves tend to take old animals in preference to those in the prime of life." However, his figures seem to indicate that the predators were selecting older animals, for of 29 wolf-killed adults, only 3 were yearlings or 2-year-olds. Burkholder (1959:7) found in his Alaskan studies that four of five ageable wolf-killed caribou adults were over 4 years old, although he states that "all of the wolf kills I could check were in excellent condition and of the 'age of primeness'."
Probably most workers would consider moose 6 to 12 years old in their prime. Indeed, the growth curve for bulls, provided by Skuncke (vide Peterson, 1955:77) shows that bulls gain weight until at least 12 years of age. Nevertheless, many of the wolf-killed moose on Isle Royale were in this category. Probably the significant point about wolf predation on any big game is not so much the selection for old animals, but rather selection against young animals (other than calves). In the present study, the invulnerability of adults less than 6 years old is striking. In addition, Murie's (1944) sample of remains of 829 Dall sheep shows that a total of only 3.4 percent of the mortality involved animals aged 2, 3, or 4 years. The figures from Tener and Stenlund, and even the limited data from Burkholder, also support the thesis that most big-game animals in their first few years are safe from wolf predation.
Young animals may be relatively invulnerable because they are more alert or they may be faster, stronger, and more agile. Perhaps they should be considered "prime," instead of the more mature individuals usually regarded as such, for security from wolves certainly is a realistic criterion. Of course, young animals could be less susceptible only because they might inhabit areas less frequented by wolves, but, at least in the case of Isle Royale moose, this seems improbable.
Incidence of Debilitating Conditions. One explanation for the vulnerability of apparently prime moose is that they are infected heavily with hydatid cysts (Echinococcus). Cowan (1947:17) proposed that the high proportion of elk kills which he found in the "prime"ages might have resulted from the heavy hydatid infections. On Isle Royale, cysts were found in the lungs of one moose each in wear-classes IV, V, and VIII. The youngest animal had only 5, but the others harbored 57 and 35, respectively. Because the parasite is contracted as moose feed, older animals should have heavier infections. Ritcey and Edwards (1958) found this to be true in British Columbia moose. In 6 or 7 years a moose might ingest enough eggs to cause a heavy infection. A very heavy infection in a moose about 4 years old was found in the British Columbia studies.
It has been well demonstrated that a higher proportion of older animals are infected with hydatid cysts than are younger ones. In Saskatchewan, Harper et al. (1955) found that only 1 of 12 immature moose examined harbored hydatid cysts whereas 13 of 31 adults were infected. The same trend was noted in deer and caribou. Rausch (1959) showed Alaska harbored cysts, but 15 percent of animals in wear-classes I, II, and III were infected, and 62 percent of those in classes VII, VIII, and IX. R. O. Skoog found cysts in only 4 of 67 caribou examined, and all 4 were over 7 years old (Rausch and Williamson, 1959).
Undoubtedly, heavily infected animals are more susceptible to predation. Fenstermacher (1937) believes that hydatid cysts are especially debilitating, and Cowan (1951:52) stated: "Infected animals are usually impoverished and of low vitality. They almost certainly have an impaired ability to survive adverse circumstances." However, Rausch (1952) disagrees, for he saw 18 cysts (80 to 90 mm. in diameter) in an Alaskan moose in apparently excellent condition. Nevertheless, it seems that such an animal would have little power to run far, or to fight off wolves. The observation by Ritcey and Edwards (quoted on page 111) is especially significant in this respect.
In Ontario, a fresh wolf-killed moose with diseased lungs was found, and signs showed that it had not fought much (Peterson, 1955). A lung was removed and sent to the Royal Ontario Museum of Zoology. Peterson (1955:176) described it as follows:
When the lung was examined in the museum laboratory it was found to be so completely filled with hydatid cysts of tapeworms that it seemed incredible that it could have functioned sufficiently to keep the animal alive during normal activity, much less allow it to ward off an attack by timber wolves. Well over 50 percent of the volume of the lung was occupied by large cysts up to one inch in diameter.
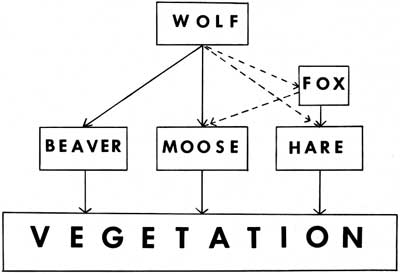
Figure 97—Most important relationships in wolf ecology. (Solid arrows indicate primary relationships.)
Crisler (1956, 1958) described a "tired" caribou running from wolves, faltering, lying down, and being killed. The lungs contained eight cysts, some as big as ping-pong balls. One of the caribou kills examined by Burkholder (1959) also harbored hydatid cysts, but the author did not elaborate.
Philosophically, it seems logical that a heavy hydatid infection should predispose a moose to predation, for the parasite depends for its perpetuation on the moose being eaten by a wolf. The greater the intermediate host's infection, the more beneficial it would be to the parasite to render the host susceptible to predation.
Old moose even without cysts or other debilitating organisms probably would be susceptible to predation because of their general infirmity and lack of agility. However, many old moose undoubtedly are infected with parasites and/or disease, which probably would increase their chances of succumbing to predation. Only two relatively intact carcasses of wolf-killed adults were examined, but each had a heavy hydatid infection. The one checked in winter also was heavily infested with ticks, and had an abnormal liver and badly congested lungs.
All that remained of most kills investigated were bones, but these also showed that many of the moose had been in poor condition. The marrow was fat-depleted or nearly so in the femurs of 5 (14 percent) of 33 adults checked; and 9 (29 percent) of 31 adult skulls or mandibles were necrotic, probably from actinomycosis (figure 86). In the total of 35 adult kills from winter plus 1 from summer, 14 (39 percent) had depleted marrow, jaw necrosis, or hydatid cysts. Some of the animals showed more than one of these conditions and probably also supported heavy tick populations (figure 84). Undoubtedly, a thorough examination of the intact carcasses of these moose would have revealed a much higher percentage of debilitating conditions.
Other studies indicate that adults of big game killed by wolves generally are old or diseased. Murie (1944) discovered that although most Dall sheep mortality occurs in the old-age category, mortality in the prime class includes a much higher percentage of diseased individuals. In the Rocky Mountain national parks of Canada, Cowan (1947) found that 17 (37 percent) of the remains of 46 adult-elk kills were either senile or diseased; and he cautioned that a condition such as hydatid infection is not evident from bones, so many others also may have been diseased. Crisler (1956:346) reported that "at least half of the kills that we observed involved crippled or sick caribou." However, all 5 fresh adult kills (4 caribou and 1 long-yearling moose) examined by Burkholder (1959) were judged to be in excellent condition, although at least one of these was infected with Echinococcus.
Species of Lesser Significance
Isle Royale wolves influence species other than moose to a much lesser degree. Some of these are less important prey animals and others are scavengers. Probably none has a great effect on the wolf, but a consideration of wolf ecology must recognize these potential and actual relationships insofar as they are known.
BEAVER
The Isle Royale beaver population apparently reached its peak from 1945 to 1950 (page 19), and in the early 1950's a sharp decline was noticed by park residents. Today beavers are common, but compared to peak numbers, the population appears low. Since wolf sign first was noticed about the time the decrease occurred, a cause and effect relationship was postulated by residents of the island.
Although beavers have not been reported as primary food for any wolf population, they supplement the diets of wolves in many areas. Beaver remains occurred in 7 percent of 420 scats from the Rocky Mountain national parks of Canada (Cowan, 1947) and composed 10.5 percent of the occurrences in 76 wolf scats from Ontario (Peterson, 1955). On Isle Royale, beavers are the second most important wolf food (table 10); beaver remains composed over 10 percent of the occurrences in 438 scats collected during the present study. The data are segregated by year and compared with previous records from Isle Royale, in table 19.
TABLE 19.—OCCURRENCE OF BEAVER REMAINS COLLECTED IN WOLF SCATS ON ISLE ROYALE
| Year | Number of scats |
Percent of occurrence |
Source |
| 1952 1954 1955 1958 1959 1960 |
87 ? 8 70 214 154 |
20 35 50 17 12 7 |
Cole, 1952a Cole, 1954 G.A. Petridesa Present study Present study Present study |
| 1958—60 | 438 | 11 | Present study |
The manner in which wolves hunt beavers is unknown, but possibly they follow shorelines until a beaver is found on land. Such an animal should be easy prey. In autumn and early spring, when beavers are most active on land, they probably are most vulnerable. For much of winter the animals are safe, but under certain conditions a few may be taken as early as March. During thaws, ice sometimes cracks along docks and islands, leaving crevices which beavers could enlarge. In 1961 when thaws occurred in late February and early March, much beaver activity was evident on March 3. Holes in the ice, trails to trees (some 100 feet from the holes) were seen along streams, islands, and docks, and in two of these places, wolves had killed beavers.

Figure 98—Beaver.
The first beaver kill was found near the northeast end of Washington Island. A trail led toward shore from a hole in the ice near a dock, and a few feet out from shore was a large blood-spattered area packed with wolf tracks. There was no sign of moose or moose remains in the vicinity. A nearby wolf was chewing what appeared to be an everted beaver hide. Two wolves were leaving the island, and four others rested about 2 miles away. We could not land and verify our aerial observations, but from the sign and the unusual behavior of the wolves, there was no doubt that they had killed a beaver.
On the same day, we found remains of a beaver killed by the pack of three near a small island in Tobin Harbor. The wolves had investigated two beaver houses situated against the island and had found trails leading from them to some fresh cuttings. A few feet from one trail there was a large bloody area covered with wolf tracks. Beaver fur was scattered about, and a well-chewed skull lay nearby.
The only literature encountered concerning predation on beaver in winter involved coyotes. Packard (1940) found tracks of a beaver that had emerged from under the ice and traveled half a mile before being killed by three coyotes. On Isle Royale, "Mr. Skadburg related the details of one incident in which coyotes killed a beaver that had been feeding upon a fallen birch from a hole in the ice" (Cole, 1952b). Extensive fieldwork in late March and April probably would produce more observations of predation on beaver.
Ever since wolves arrived on Isle Royale, they undoubtedly have been killing beavers, but whether wolves are directly responsible for the beaver decline is unknown. Before the wolf population was established, many colonies had been abandoned (Krefting, 1963), and supplies of aspen, the preferred food, in the southwest and central sections of the island were judged low (Krefting, 1951). There is a reserve of white birch on most of the island even today, and beavers certainly eat birch, but whether they can thrive on it has not been determined.
T h e following statements by Cowan (1947:169) indicate that, at least in his study area, wolves were not primarily responsible for a beaver decline:
In 1930 the Athabaska Valley of Jasper Park was superlative beaver range and bore a very heavy population despite the abundance of predatory animals. The removal of trees by these beavers at the time when the elk and moose were eliminating all seedling trees has had the inevitable result of rendering large areas of the park unsuitable for beaver.
There is little doubt that wolves and other predators are effective in reducing a beaver population that has eaten itself out.
I have seen no evidence that predators can prevent a beaver population from increasing until it is so large that safely available food becomes inadequate.

Figure 99—Snowshoe hare.
On Isle Royale, beavers that had depleted their aspen supply probably moved to new sites or traveled long distances to obtain aspen, increasingly exposing them to predation. In such a case, predation might be the immediate cause of a beaver decline, but the basic cause would be the shortage of preferred food. Undoubtedly any adverse conditions undergone by a beaver population will result in increased predation; the important distinction to make is whether wolves are primary or secondary causes of the decline. Little work on this aspect was done during the present study, but research is now underway on the ecology of Isle Royale beavers. It should furnish basic information from which a more definite statement can be made about wolf-beaver relations.
SNOWSHOE HARE
The Isle Royale hare population conceivably is a potential food source for the wolves. In British Columbia, Stanwell-Fletcher (1942:139) found that when the snow crusted ". . . snowshoe rabbits became once more a part of the wolf diet." Murie (1944:58) maintained that "if hares were plentiful they would probably supplement the food supply of wolves considerably." However, it appears that, on Isle Royale, hares are eaten only incidentally and that even a high population hardly would affect the wolf.
Isle Royale hare numbers increased considerably during the present study, as evidenced by increased percentages of hare remains in fox scats, reports from island residents, and observations of hares and tracks (table 20). Even so, the percentage of occurrence of hare remains in 438 wolf scats was only 3.1 percent, and no increase was evident from 1958 to 1960. The low representation of hare is at variance with findings by Cole (1952a) that hare remains composed 24 percent of the winter food of Isle Royale wolves, based on 87 scats. However, since coyotes still were present in 1952, possibly some scats were from coyotes, for Thompson (1952) demonstrated a great overlap in scat sizes of these species. The present figures do agree with statistics by Cowan (1947) and Thompson (1952), who reported that hare remains composed 7 percent of 420 scats from the Rocky Mountain national parks of Canada, and 5 percent of 435 scats from Wisconsin, respectively.
TABLE 20.—EVIDENCE OF INCREASE IN HARE NUMBERS
| Year | Percent of occurrence of hare remains fox scats |
Weeks of summer fieldwork per hare observation |
Hours of flying per hare observationa |
| Summer 1958 | 17 | 7.0 | ... |
| Winter 1959 | ... | ... | 115 |
| Summer 1959 | 46 | 5.0 | ... |
| Winter 1960 | ... | ... | 46 |
| Summer 1960 | 52 | .7 | ... |
| Winter 1961 | ... | ... | 22 |
a Seen from aircraft during moose census or observations of wolves hunting.
Observations of wolf-hare encounters also indicate that hares are not important to Isle Royale wolves. On February 6, 1961, the pack of 15 accidentally flushed three hares but paid no attention. Even a lone wolf, hunting moose, showed no interest in three hares it flushed, on February 3, 1961. In one of these instances, a hare circled counterclockwise and passed in front of the wolf; 20 seconds later the wolf crossed its trail without investigating.
To substitute enough hares for the average of 10 to 13 pounds of moose consumed daily, each wolf would have to take two or three a day, which might be an arduous task. Wolves probably have more difficulty catching hares than do foxes (which feed regularly on them), because the latter can follow hares into thicker cover and smaller hiding places, and over deep snowdrifts. It probably is more efficient for wolves to concentrate on moose.
RED FOX
Foxes seem to be common on Isle Royale but not abundant. A strong limitation on numbers probably is winter food supply, for mice are relatively unavailable, and at least during the first year of the study, hares were scarce. Information on the diet of the fox was obtained by analysis of 295 scats collected from trails during three summers (table 21). Probably the sample was biased toward winter, for summer scats containing berries disintegrate quickly. Nevertheless, the dependence of foxes on hares, at least during winter, is evident. Since the hare population is growing, perhaps fox numbers also will increase.
TABLE 21.—ANALYSIS OF FOOD REMAINS IN 295 FOX SCATS COLLECTED FROM TRAILS
| 1958 | 1959 | 1960 | Total | |
| Number of scats Number of occurrences |
20 34 |
113 152 | 162 191 |
295 377 |
| Food items | Percent of occurrence | |||
|
Snowshoe hare Bird Muskrat Moose Insect Mouse Red squirrel Snake Unidentified mammal Strawberry Shadberry Miscellaneous a Unidentified fruit Unidentified |
17 .. 6 6 17 9 3 9 .. 12 9 .. 12 .. |
46 10 11 6 7 2 5 5 3 1 .. 3 .. .. |
52 15 8 6 .. 3 2 2 5 1 2 2 .. 2 |
47 12 9 6 4 3 3 3 3 2 2 3 1 1 |
It is interesting that the most foxes observed from the air per day (10, on February 27) were seen in 1961. This is six more than the maximum per day seen during the first two winters. Although complete figures were not kept on number of animals observed per study period, until 1961, the highest number (29) definitely was seen in 1961. However, conclusions on population changes should not be based on observations alone, for biases are many. Most foxes are seen at moose remains on lakes, bays, and shores, where they are most apparent from the air. Variations in amount of time spent over any of these locations could produce widely different totals of animals seen. Since foxes were studied only incidentally, no evidence was obtained that allows absolute, or even relative, estimates of population size.
 Figure 100—Red fox. Photo by Don Murray. |
The most important relationship between the fox and the wolf involves the food gleaned by foxes from remains of wolf-killed moose. Although moose remains composed only 6 percent of the occurrences in the fox scats collected, a higher percentage of the winter diet may consist of moose. Scats on trails (the only places where scats were collected) would be left primarily by traveling animals, but many foxes apparently remain near moose kills for days; most of their scats would not be represented in the collection. During winter, foxes feed on almost every kill soon after the wolves leave, and on one occasion four animals were seen feeding on a carcass.
Although wolves generally benefit foxes, they sometimes kill them. Twice, foxes were seen to flee instantly upon sensing wolves. Another time, on March 15, 1960, I watched a wolf kill a fox. About 5:05 p.m. the large pack was heading through a spruce swamp about a mile southwest of Halloran Lake when suddenly the lead animal sprang toward a running fox 125 yards away. As the wolf passed a moose carcass, from which the animal had run, a second fox scurried off. Within about 15 yards the wolf caught the fox and shook it violently. It then carried the limp carcass under some trees. Half an hour later, I found that the wolf had ripped out the intestines of the fox and abandoned the animal, at least temporarily. The next day the carcass was gone; it may have been eaten or just carried back under the trees.
A contrasting observation was made on the day after the fox was killed. While the wolves fed on the moose carcass, a fox lay curled up about 100 feet away, apparently fast asleep. On another occasion, a fox ventured to within 100 feet of a lone wolf feeding on a moose carcass. Cole (1957) twice saw foxes closely approach wolves near moose carcasses, and found that "they seemed to have little fear of the larger animals when abundant supplies of food existed nearby." Murie (1944) also reported instances in which foxes showed no fear of wolves. He concluded, in his Mount McKinley study, that "the relationship between the wolf and the fox seems to be one of mutual gain." Foxes secure food from wolf kills, and wolves enlarge fox dens for their own use.
On Isle Royale six fox pups were produced in 1961 in a den which certainly was large enough for a wolf den (page 71 and figure 60). In the previous 2 years it was vacant, and its original occupants remain unknown. This was the only den found, so nothing is known about this aspect of wolf-fox relationships on Isle Royale.
RAVEN
A peculiar relationship exists between the large pack of wolves and a flock of ravens. At every fresh kill, ravens perched in nearby trees, waiting to feed. The instant the wolves finished eating, these scavengers alighted on the carcass. Apparently kill remains provide the primary winter food for Isle Royale ravens.
Probably because these birds are so dependent on the wolves, small flocks regularly accompany the animals during their travels. They fly ahead of the pack, perch in trees until the wolves pass, and then "leap frog" them again. Frequently, I even saw ravens tracking wolves. A bird so engaged flies directly over a string of tracks. Upon discovering a wolf scat, it thoroughly picks this apart and presumably swallows all edible portions, and then continues along the trail. I cannot remember any raven backtracking, so perhaps the birds deliberately track the wolves to overtake them and only feast on scats incidentally.
Once while the wolves attacked a moose, the ravens swirled around them excitedly. After the wolves wounded the moose, one bird sat in a tree and cawed as they tried to make the moose run. Sometimes the scavengers joined wolves in eating bloody snow.
Wolves and ravens often seem to play together, especially when the wolves rest on the ice, fully gorged (figure 101). The following account of activity noted on March 5, 1961, includes the range of "playful" behavior witnessed between wolves and ravens. As the pack traveled across a harbor, a few wolves lingered to rest, and four or five accompanying ravens began to pester them. The birds would dive at a wolf's head or tail, and the wolf would duck and then leap at them. Sometimes the ravens chased the wolves, flying just above their heads, and once, a raven waddled to a resting wolf, pecked its tail, and jumped aside as the wolf snapped at it. When the wolf retaliated by stalking the raven, the bird allowed it within a foot before arising. Then it landed a few feet beyond the wolf, and repeated the prank.

Figure 101—Ravens about to tease some resting wolves.
Crisler (1958) who observed similar activity from the ground, described it as follows:
He [a raven] let the pups trot to within six feet of him, then rose and settled a few feet away to await them again. He played this raven tag for ten minutes at a time. If the wolves ever tired of it, he sat squawking till they came over to him again.
Although Isle Royale wolves almost caught teasing ravens several times, I never saw them succeed; neither were raven remains found in any of the 438 wolf scats analyzed. Therefore, it appears that either the ravens are thoroughly familiar with the wolf's capabilities, or the wolves do not seriously attempt to capture the ravens.
COYOTE
Coyotes were present on Isle Royale from the early 1900's, but by February 1957, very few remained (page 19). Since no coyote or coyote sign was found during the present study, undoubtedly the species has been extirpated from Isle Royale. The cause of this is unknown, but the wolf may be responsible. During a study of British Columbia fauna, Munro (1947) recorded a report from a native who had found remains of a coyote killed and eaten by wolves. The man believed that where wolves invade an area, they drive the coyotes out. Minnesota wardens also discovered a coyote killed by wolves (Stenlund, 1955:46): "The male coyote had run onto the lake from the woods and was immediately killed by the wolves [three] which were running on the ice." Since coyotes and wolves are closely related and since wolves are strongly territorial, it is not unlikely that on a limited range, such as Isle Royale, wolves would chase, and probably kill, every coyote encountered.
| <<< Previous | <<< Contents>>> | Next >>> |
fauna/7/sec4.htm
Last Updated: 01-Feb-2016