|
NATIONAL PARK SERVICE
Giant Sequoia Ecology: Fire and Reproduction |

|
CHAPTER 1:
INTRODUCTION
Howard S. Shellhammer
Giant sequoias [Sequoiadendron giganteum (Lindl.) Buchholz] are among the most magnificent living things on earth. They are gigantic in stze, beautiful to behold, awe inspiring, and have had a long evolutionary history. Once discovered by western man, the giant sequoias were quickly threatened by the lumberjack's saw and axe. Protection came only under the auspices of federal, state and local governments. This protection, however, has been so complete as to preclude the very natural forces which are required for the perpetuation of the species. The absence of fire from the sequoia groves, for example, has created a condition where the subtle, yet relentless, process of ecological succession and the violent force of potential crown fires again threaten the species. Enlightened management is called for to insure their continued existence.
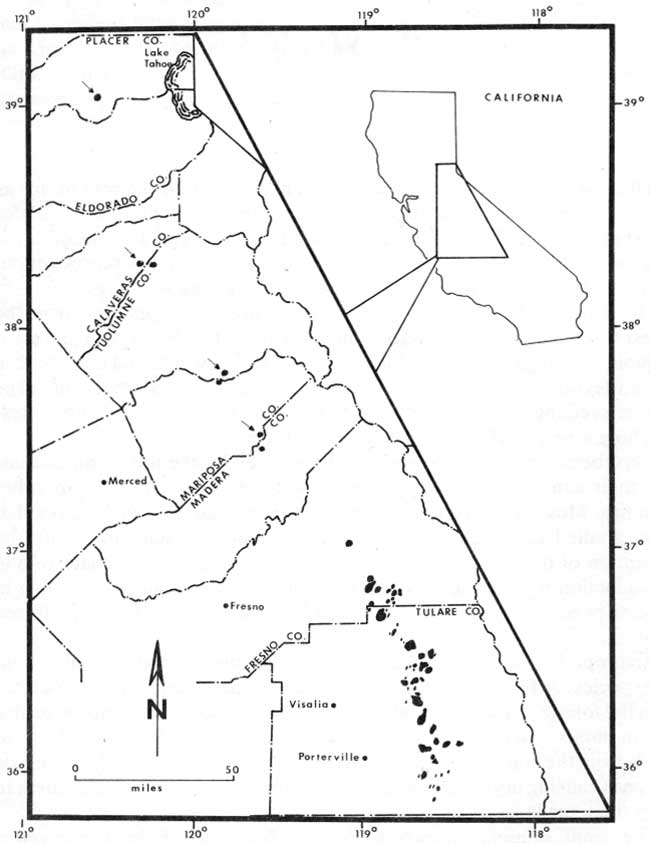
Giant sequoias have a disjunct natural distribution. They are restricted to a series of relatively distinct groves which extend some 420 kilometers (260 miles) along the west side of the Sierra Nevada (Fig. 1). Rundel (1972b) listed seventy-five community units or groves, but there is considerable controversy over what constitutes a grove (Hartesveldt et al. 1975). Giant sequoia groves are mainly restricted to elevations between 1525 and 2290 meters (5,000 to 7,500 feet) but may be found growing naturally as low as 1,220 meters (2,700 feet) or as high as 2,713 meters (8,900 feet). Most of the groves are located in the southern third of their geographical range and in or near Sequoia and Kings Canyon National Parks (Fig. 2). While the groves vary from less than 1 to 1,619 hectares (1 to 4,000 acres) in extent, nowhere does the giant sequoia exist as an extensive pure stand. They are associated with those trees which Munz and Keck (1959) have described as comprising the yellow pine forest community: white fir (Abies concolor), sugar pine (Pinus lambertiana), Ponderosa pine (Pinus ponderosa), incense cedar (Calocedrus decurrens), black oak (Quercus kelloggii), and often Douglas fir (Pseudotsuga menziesii), red fir (Abies magnifica) and/or Jeffrey pine (Pinus jeffreyi). The particular combination and frequency of these trees in a given grove depends on elevation, latitude, exposure, soil moisture, and the length of time it has been afforded protection from lumbering and fire. Rundel (1969) has pointed out that there is no other species of higher plant found only in association with giant sequoias. Hence, even though giant sequoias are clearly dominant in the mixed coniferous forests in which they occur, they cannot be considered to constitute a distinct plant community in themselves, for as Dice (1952) has suggested, a plant community should have at least two species in common.
|
| Fig. 1. The range of the giant sequoia (adapted from a map by the California Department of Natural Resources). (click on image for an enlargement in a new window) |
Giant sequoias were more widely spread and probably occurred in continuous forests in the past. The earliest close relatives of the present species, Sequoia reichenbachi and S. couttsiae, were distributed through out Greenland, Alaska, Canada, and England during Cretaceous and Tertiary times (Chaney 1951). Fossil sequoias considered to be directly ancestral to the present giant sequoia are found in Miocene deposits in Idaho (Axelrod 1959, 1964). A more recent direct ancestor, Sequoiadendron chaneyi, has been described by Axelrod (1962) from a Pleistocene forest in western Nevada. Giant sequoias moved southwestwardly during the late Tertiary as they vanished from the rest of their former range; S. chaneyi disappearing from Europe as late as the Peistocene (Martin 1957). Axelrod (1964) has proposed that the southwesterly migration from Idaho to California was induced by the changes in climate accompanying the gradual rise of the Sierra Nevada. Winter temperatures became more severe in the north, and summer precipitation decreased in the east during the time of migration (Wilson 1928). Hartesveldt et al. (1975) postulated that the giant sequoias crossed the Sierra Nevada in several places and became established on the western slope long before the mountains attained their present height.

|
| Fig. 2. A portion of the Giant Forest Grove in the Crescent Meadow area of Sequoia National Park. |
Numerous hypotheses have been offered to explain the present, narrow, disjunct distribution of giant sequoias. Muir (1876), Stark (1968a, 1968b), Wulff, Lyons and Dudley (1911), and Shinn (1889) all suggested that giant sequoias have a narrow range of tolerance to climatic factors. Most of these authors, especially Muir, envisioned glaciers and cold air drainage as possible factors in disrupting the postulated continuous distributional pattern. Winter cold has often been mentioned, but is poorly understood as a potential factor in limiting growth (Hartesveldt et al. 1975). Soil apparently plays a minor role as a controlling factor, in as much as the parent rock material varies considerably between groves. Giant sequoias have grown vigorously when planted in diverse soils throughout the world (Hartesveldt et al. 1975). Rundel (1969, 1971, 1972a) proposed that the present boundaries of the groves are controlled by an interaction of soil moisture, temperature and the ecological tolerance of the seedling stage. We adhere to this view, as do most students of giant sequoia ecology at the present time.
Giant sequoias were first discovered by western man in 1833 and again in 1852, when their "discovery" was more widely publicized (Hartesveldt et al. 1975). Lumbering began during 1852, and whole groves were being cut down by 1856. The first efforts to preserve some of the groves began soon after their second discovery in 1852. The Mariposa Grove, which is now part of Yosemite National Park, was deeded to the State of California by the federal government in 1864 to be administered as a state park. Additional groves were protected with the establishment of Yosemite, Sequoia and General Grant National Parks in 1890. Other groves were given protection by federal, state and private actions between 1890 and 1940. The last remaining large tract of unprotected sequoia land was added in 1940. In that year the Redwood Mountain Grove, measuring 1,506 hectares (3,720 acres), was incorporated into the newly created Kings Canyon National Park. At present more than 92% of all sequoia grove acreage is under public ownership.
Lumbering continued sporadically until the mid-1950s, by which time approximately 34% of the original total acreage had been cut. Current policies of the various governmental agencies prohibit lumbering of giant sequoias on public lands, and little lumbering of this species is conducted on the remaining private lands. Since fire suppression also has been a general policy of the controlling agencies, most groves have been rigidly protected from fire. Although they have received a full measure of protection, the trees have been the subject of but a few research investigations. The recognition of the need for more sound information regarding these trees and their requirements for survival provided the basis for the current study. Findings of these investigations are reported in this publication.
The management of giant sequoias was one of passive preservation until approximately ten years ago. The change to an active, research-supported management plan, especially on the part of the National Park Service, was stimulated by a series of events. A considerable body of evidence was accumulating on the ecological role of fire in the Sierra Nevada in general and in giant sequoia groves in particular. Scientists were reporting, as well, on the changes taking place in the protected groves, both in the nature of human impact and the increased fire danger from increasingly dense vegetation and fuel accumulation. Finally, the Leopold report of 1963 urged that biotic associations in each national park be maintained, or where necessary recreated, as nearly as possible in the condition that prevailed when the areas were first seen by European men.
The role of fire was noted as early as 1878 by John Muir. Mason (1948) noted that sugar pines decreased in number in areas devoid of fires for an extended period of time. Both Biswell (1961) and Hartesveldt (1963, 1964) suggested that fires burned through Sierran communities with a degree of regularity before the advent of western man. Hartesveldt (1963) documented the occurrence of fires in the Mariposa Grove of Yosemite National Park by analyzing cores taken from giant sequoias. He and other scientists postulated that frequent surface fires maintained a more open forest prior to the arrival of European man (Weaver 1947, 1951, 1964; Biswell 1967; Hartesveldt 1963; Hartesveldt and Harvey 1967; and Kilgore 1971, 197-2). Show and Kotok (1924) suggested, however, that crown fires did not accompany surface fires as the former were virtually nonexistent in the Sierra Nevada during pristine times, and this hypothesis has been supported by Kilgore and Sando (1975). Kilgore (1973) summarized the present outlook on the role of fire in a mixed-conifer forest. He stated that "fire (1) prepares a seedbed, (2) cycles nutrients within the system, (3) adjusts the successional pattern in various ways, (4) modifies conditions that favor wildlife, (5) influences the mosaic of age classes and vegetation types, (6) alters numbers of trees susceptible to attack by insects and disease, and (7) both reduces and creates fire hazards." He further recognized that each of these functions is affected by fire intensity and frequency. Most of these functions will be discussed in various parts of the present monograph.
The studies of Hartesveldt (1962, 1964) on the effect of human impact on the giant sequoias of the Mariposa Grove of Yosemite National Park provided the basis for a change in attitude toward grove management. He suggested that trampling about giant sequoias may increase, rather than decrease, their rate of growth by reducing other plants which compete for soil nutrients and moisture. He and Beetham (1962) and later Stark (nee Beetham 1968a, 1968b), demonstrated that the absence of sequoia reproduction under protected conditions is due to the fact that young giant sequoias are intolerant of shade. Giant sequoias are favored in earlier stages of succession which may be created when fire disrupts the climax community. Each author showed that even though germination occurs under shaded conditions, seedling survival is severely curtailed. Both reported that seedlings survive best in bare mineral soil, but that the seedlings have very little chance of becoming established when the seeds germinated on top of a thick layer of litter and duff that accumulates in the absence of periodic fire. Hartesveldt (1964) concluded that the advanced plant succession found in present-day groves is increasing the opportunity for large crown fires which both he and Show and Kotok (1924) determined were absent in pristine times.
Finally, the Leopold report (1963) was important in creating a "climate" for a new approach to park management. Earlier, Olmstead (1952) and Wright (1933) had both urged broad ecological studies be conducted to provide the basis for park management. Hartesveldt (1962, 1964) recognized the need for further research on the giant sequoia. The Leopold Report (1963), which was developed by a special committee appointed by Secretary of the Interior Udall, stressed the ecological complexity of the national parks and implied that a more thorough understanding of each area was necessary in order to effectively manage them. The present study has been carried out with this philosophy in mind. It is an attempt to provide some of the basic information necessary to the scientific management of sequoia groves.
Giant sequoias have long been associated with fire (Hartesveldt 1964). They possess many ecological adaptations to fire. They have a thick bark which helps to insulate them from heat. They shed enormous numbers of seeds which have the greatest chance of success when they fall on soil cleared by fire or erosion. The seedlings are intolerant to shade. They are often overgrown by shade-tolerant plants which crowd in as succession proceeds and which generally increase the chances of the needed surface fires. But fire, it should be noted, is but one part of the ecology of these trees.
Our intent, in the light of the factors previously discussed, has been to study the general ecology of the giant sequoia so that the role of fire and other factors may be more completely understood, and to provide information necessary for the development and implementation of an ecologically sound resource management plan.
Holloway (1954) said it best when he suggested that studies of fire ecology must be "founded in a deep appreciation of the forest, of its complex interrelationships of all its component parts. And, since the forest is an ever-changing, living community, understanding also demands unrelenting study of all trends in forest evolution, man-made or natural. For a single forest, the complex actions and interactions of all plants, soil, animals, microorganisms, and man together with all factors of climate, topography, lithology, and history must be studied together and in synthesis."
| <<< Previous | <<< Contents >>> | Next >>> |
chap1.htm
Last Updated: 06-Mar-2007