|
NATIONAL PARK SERVICE
Giant Sequoia Ecology: Fire and Reproduction |

|
CHAPTER 4:
VEGETATIONAL CHANGES
H. Thomas Harvey
Introduction
Secondary succession may occur in forest ecosystems after a disturbance such as fire. This succession may be assessed by measuring changes in species composition and their abundance (Horn 1974). The succession we were primarily concerned with followed experimental treatments consisting of mechanical and manual disturbances and fires. The fires were either surface fires or burn pile fires.
The vegetational changes that were monitored consisted of ground vegetation (higher plants up to a meter tall) and trees (single-stemmed woody plants more than a meter tall).
Bonnicksen (1975) has studied the mixed conifer—giant sequoia forest ecosystem in Redwood Canyon immediately northeast of our study areas. He concludes that the primeval forest "consisted of a mosaic pattern of gaps, and approximately even-aged groups of trees and shrubs in various stages of development." He also presented a successional pathways model in which the sequences progress toward a multilayered vegetation unit of pure white fir. However, Bonnicksen suggests that fire, insects, disease, and windthrow probably prevented most of the forest's progressions from reaching the white fir climax stage. Thus the sequoia—mixed conifer forest is more a property of the mosaic than of any single vegetation unit. He concludes that this forest ecosystem consists of "steady state space-time mosaics composed of distinct structurally defined, and successionally related, vegetation units." In other words, small areas of different species at different stages of development form a mosaic.
Concern has been expressed by several investigators (Biswell 1961; Hartesveldt 1964; Rundel 1971) that the absences of fire may eventually lead to the succession of white fir in giant sequoia communities, and thus place the relict species, giant sequoia, in jeopardy (Fig. 16.).

|
| Fig. 16. White fir saplings growing beneath giant sequoias. |
One basic objective of these studies was to determine if certain early stage species of low occurrence in the present community would increase in number upon introduction of fire and manipulation, while other more common species might decrease after treatment. It was also an objective of the study to document the rate at which the return to pre-burn conditions was accomplished. The population fluctuations of all the species were monitored in both the treated sections and the controls in order to ascertain the relative effects of the manipulation and fire.
An inventory of tree species was taken in both treated and control sections in addition to the surveys of ground species of herbs and shrubs. Particular interest centered on white firs and giant sequoias, as they are respectively considered to be climax and disclimax dominants in those areas where physical factors permit giant sequoias to grow. Harlow and Harrar (1969) listed the tree species that occur in the study areas with respect to the conditions of shade tolerance as follows: giant sequoia and ponderosa pine, intolerant; sugar pine and incense cedar, intermediate or cedar may also be considered to be tolerant; and white fir, tolerant. It was hypothesized that if fire favors giant sequoias they should show relatively better survival and growth than white firs in the manipulated sections in comparison to the control sections.
Methods and materials
In order to assess vegetational changes following manipulation, a series of permanent plots was established in the study areas (Mueller-Dombois and Ellenberg 1974). For ground vegetation analysis 2 m2 plots were established in both treated and control areas. They were surveyed as to number and species of plants less than a meter in height prior to treatment, one year after treatment, three or four years after, and eight or nine years after treatment. A one meter square grid marked off in square decimeters was used to facilitate location and counting of plant species (Fig. 17).

|
| Fig. 17. Meter square grid being used by Tom Harvey. |
Permanent regularized plots measuring 5 m x 20 m were established in three areas for assessing tree vegetation greater than 1 m in height. These plots were examined prior to manipulation, and the treated sections were surveyed one year after manipulation and fire, and both types of sections were surveyed again in 1974. In addition tree populations were assessed by the random pairs method (Phillips 1959) in all but North Area. Trees were selected and measured as to distance apart, species and diameter at breast height. In 1974 a complete survey of snags was made in all study areas, and a comparison made as to species, size, and abundance in treated versus control sections.
Results
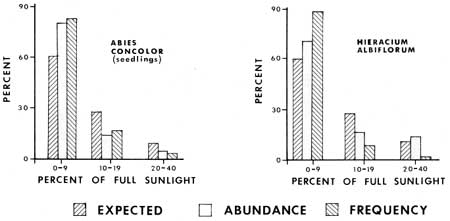
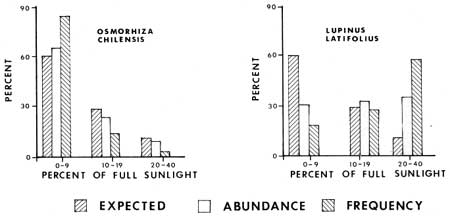
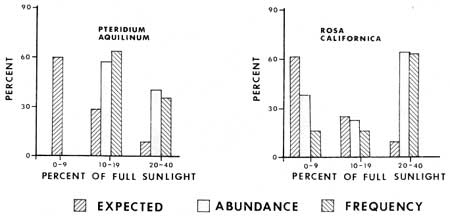
Light determinations were made at 114 2 m2 ground vegetation plots, thus making an analysis of levels of relative capacity to tolerate shade by some plants possible (Figs. 18-20). Each bar representing percent expected was determined by dividing the total number of sample plots into the number of plots that had a given light value; e.g. 60% of the sample plots had sunlight values of 0-9%. The abundance and frequency percentages of various species are based on the relative occurrence of a species population in plots with given light values. For example, of all the white firs in sample plots, 82% are in plots with 0-9% of full sunlight. Therefore one can contrast what might be expected if a plant was distributed in dependently of sunlight values with its actual distribution. From the graphs (Figs. 18 and 19) it is evident that Abies concolor, Hieracium albiflorum and Osmorhiza chilensis had higher frequencies and levels of abundance in the low categories than there were plots with those light intensities. Galium triflorum, Pyrola picta and Adenocaulon bicolor had similar responses, with more than 20% higher values for abundance and/or frequency in the 0-9% full sunlight category.
|
| Fig. 18. Plant response in 114 plots to percentage of full sunlight (explanation in text). |
|
| Fig. 19. Plant response in 114 plots to percentage of full sunlight (explanation in text). |
|
| Fig. 20. Plant response in 114 plots to percentage of full sunlight (explanation in text). |
Lupinus latifolius, Pteridium aquilinum and Rosa californica were under-represented in the low light plots and over-represented in the higher lighted plots (10-40% full sunlight). Two plant species, Galium sparsiflorum and Viola lobata were more intermediate in their occurrence with respect to light levels. It is of interest to note that the two Galium species occur in the same habitat, but that the species with more leaves at the nodes (G. triflorum with six leaves) is apparently more shade tolerant than the species with fewer leaves (G. sparsiflorum with four leaves).
Light determinations were made in the dense stand of giant sequoia seedlings in Trail Area by randomly placing ten test vials at the site. An average of 13.6% of full sunlight was obtained with a range in values of 10% to 16%. Measurements were also made at locations of individual sequoias, and a correlation coefficient calculated with respect to height. The results are discussed in Chapter 5.
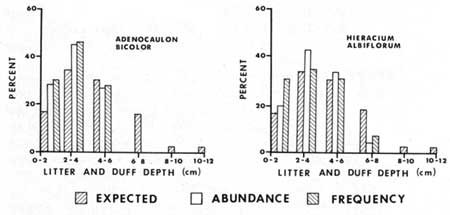
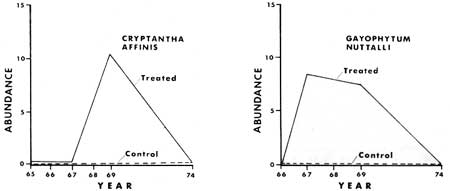
Most (64%) of the litter and duff plots were in the 2-6 cm deep categories. No plants were found in litter and duff greater than 8 cm (3.2 in) deep, however the sample size was too small to draw any inferences. All but Abies concolor had percent frequency of occurrence less than expected in the 6-8 cm class (Figs. 21-23). Expected frequency and abundance percentages were calculated in the same manner as for light described above. Adenocaulon bicolor, Hieracium albiflorum, Osmorhiza chilensis, and Pteridium aquilinum had higher than expected percentages of their abundance and/or frequency in the thinner duff-litter classes, i.e. in 0-2 cm and 2-4 cm classes. Both Abies concolor and Pyrola picta appear to be able to tolerate the deeper litter-duff thickness (Fig. 23), as they were at or below the expected percentage in the 0-2 cm class and not at or above the expected percentage in the 4-6 cm class. The annuals Cryptantha affinis, Gayophytum nuttallii and Montia gypsophiloides were found only in the 0-2cm litter-duff class, which implies an intolerance to thick duff.
|
| Fig. 21. Plant growth versus litter and duff thickness (explanation in text). |
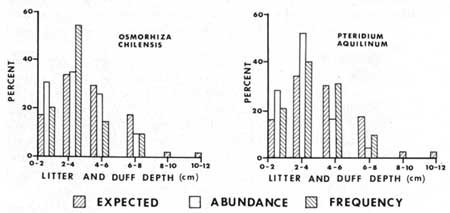
|
| Fig. 22. Plant growth versus litter and duff thickness (explanation in text). |

|
| Fig. 23. Plant growth versus litter and duff thickness (explanation in text). |
The ground vegetation in the different study areas varied slightly in the ranking of abundance and frequency of occurrence of the various species present. It also showed minor variation from year to year, especially in the treated sections. A listing of the treated section in one of the areas (North Area) prior to treatment indicates the relative importance of most of the plant species (Table 4). A listing of all plant species observed in the study areas is in Appendix I. Voucher specimens are in the San Jose State University Herbarium.
Table. 4. Relative importance of ground plant species in North Area.
| Species | Relative Abundance (% of total ind.) |
% Frequency (% of total) | Mean Density (No./m2) |
| Lupinus latifolius var. columbianus | 20 | 50.0 | 2.4 |
| Hieracium albiflorum | 15 | 42.8 | 1.9 |
| Viola lobata | 13 | 71.4 | 1.6 |
| Rosa californica | 11 | 32.1 | 1.4 |
| Osmorhiza chilensis | 8 | 46.4 | 0.9 |
| Silene bridgesii | 4 | 3.5 | 0.5 |
| Pyrola picta | 3 | 25.0 | 0.4 |
| Pteridium aquilinum | 3 | 14.2 | 0.4 |
| Galium triflorum | 2 | 10.7 | 0.3 |
| Viola purpurea | 2 | 7.1 | 0.3 |
| Adenocaulon bicolor | 1 | 14.2 | 0.1 |
| Gayophytum Nuttallii | 1 | 3.5 | 0.1 |
| Smilacina racemosa var. amplexicaulis | 1 | 10.7 | 0.1 |
| Viola Sheltonii | 1 | 7.1 | 0.1 |
| Potentilla glandulosa | 1 | 7.1 | 0.1 |
| Castanopsis sempervirens | 1 | 3.5 | 0.1 |
| Draperia systyla | 1 | 3.5 | 0.1 |
| Disporum trachyandrum | 1 | 3.5 | 0.1 |
| Goodyera oblongifolia | 1 | 3.5 | 0.1 |
| Habenaria unalascensis | 1 | 3.5 | 0.1 |
| Phacelia nutabilis | 1 | 3.5 | 0.1 |
| Rubus leucodermis | 1 | 3.5 | 0.1 |
| Ribes Roezlii | 1 | 3.5 | 0.1 |
| Symphoricarpos mollis | 1 | 3.5 | 0.1 |
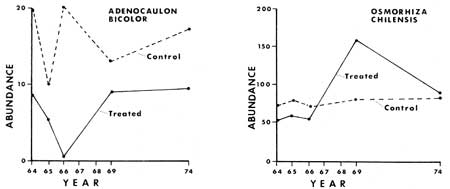
Some of the results from the monitoring of the ground cover vegetation (tracheophytes less than 1 m tall) before and after manipulation are presented graphically in Figs. 24-33. From these graphs it is apparent that certain species show similar response patterns. For example, several annuals that increased significantly following treatment were practically gone seven to ten years after manipulation. Specifically, Cryptantha affinis, Gayophytum nuttallii and Gilia giliodes were never found in the control plots of any of the areas, but increased from zero in number in the manipulated plots of North and Ridge Areas the year or two after manipulation. They then disappeared, or, as in the case of G. giliodes, were reduced in number during the seven years after the treatment. The other two study areas (Trail and Ridge) were apparently unfavorable to growth of annuals, possibly being either too dry or having an inadequate seed source.
Certain perennial and biennial plants showed a pattern of population numbers similar to the annuals. They were lacking in the control section plots throughout the study and treatment plots prior to disturbance. In the fire or manipulated plots both types increased dramatically following manipulation or fire, and then gradually decreased. This was especially well documented in the case of Ribes roezlii. For example, 12 plants were present in the 10 2 m2 treated plots of Ridge Area a year after the fire, while nine years later only one plant had survived. Larger populations of R. roezlii in the other study areas showed a similar pattern (Figs. 25, 30). Phacelia mutabilis responded in a similar fashion (Figs. 26, 31). Other species responded in various ways, as will be discussed below.
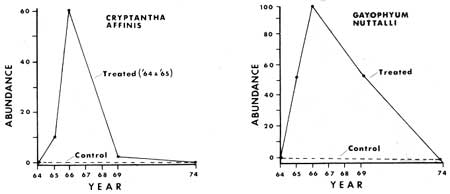
|
| Fig. 24. Plant numbers in 52 2 m2 plots in response to treatment in North Area in 1964 and 1965. |
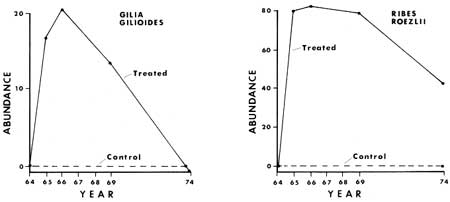
|
| Fig. 25. Plant numbers in 52 2 m2 plots in response to treatment in North Area in 1964 and 1965. |
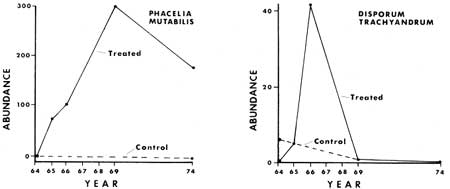
|
| Fig. 26. Plant numbers in 52 2 m2 plots in response to treatment in North Area in 1964 and 1965. |
The genus Ceanothus was represented by only a few individuals in the study areas prior to manipulation and fire. Three species were present, namely C. integerrimus, C. parvifolius and C. cordulatus. Only the first two species occurred in sampling plots in South Area. At this lower elevation site C. integerrimus increased from one during pretreatment to 207 in 1974 in the same plots. C. parvifolius increased in the same interval from three individuals to a sample population of 1399, and a 50% frequency level. C. parvifolius was also observed growing on the perimeters of the burn piles in North Area in the summer of 1965 after the piles had been burned the previous September. A total of twenty-seven plants were observed at the edges of 10 of the 26 burn piles.
The apparent response to treatment of Lupinus latifolius var. columbianus was of special interest in North Area. Prior to manipulation in 1964 this species covered approximately 40% of the section to be treated. Ten years after fire and manipulation the lupine cover was about 90%.
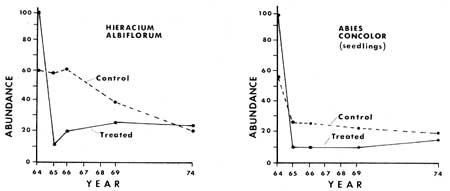
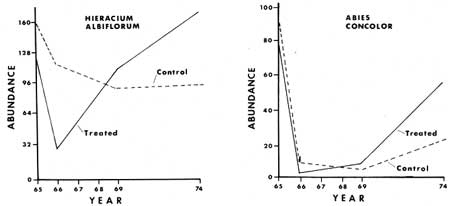
The fluctuations of those plant species already in fair number in the areas prior to fire and manipulation were less clear in the response after treatment. The three species of greatest abundance were Hieracium albiflorum, Adenocaulon bicolor and Osmorhiza chilensis (Figs. 27, 28, 33). Hieracium albiflorum apparently responded by a rapid decrease the year after treatment, but quickly recovered, and in four or five years had returned to population numbers equal to those of pretreatment conditions. Osmorhiza chilensis remained at essentially equal levels in the control and treated sections in North Area (Fig. 28). However, in the other three areas there was a significant decrease of populations in the manipulated plots a year after manipulation, as they dropped by about 72%, while control populations only dropped about 22% in abundance. Total population in the control plots was 462, which dropped to 361, while in the treated plots the total population dropped from 515 to 147 after treatment. At the last assessment in 1974 the total inventories were very similar between treated and control plots.
|
| Fig. 27. Plant numbers in 52 2 m2 plots in response to treatment in North Area in 1964 and 1965. |
|
| Fig. 28. Plant numbers in 52 2 m2 plots in response to treatment in North Area in 1964 and 1965. |
|
| Fig. 29. Plant numbers in 37 20 m x 1 cm transects in response to treatment in South Area in 1966. |

|
| Fig. 30. Plant numbers in 37 20 m x 1 cm transects in response to treatment in South Area in 1966. |

|
| Fig. 31. Plant numbers in 37 20 m x 1 cm transects in response to treatment in South Area in 1966. |

|
| Fig. 32. Plant numbers in 41 2 m2 plots in response to treatment in Trail Area in 1965. |
|
| Fig. 33. Plant numbers in 41 2 m2 plots in response to treatment in Trail Area in 1965. |
Tree vegetation over 1 m (3.2 ft) in height was found to consist mainly of white fir. Data combined from all four areas indicate that 76.5% of all conifers were white fir, 13.7% were sugar pines, 6.6% were giant sequoias, and 3.0% were incense cedars. In North Area the population of white firs in the treated section was reduced 93%, from 1,010 to 66 per hectare, after 10 years. During the same period the control section had apparently decreased by 21%, from 1,030 to 811 per hectare. The density of white firs was essentially the same for the treated and untreated sections prior to treatment, i.e. 1010 and 1030 per hectare respectively.
In the complete survey of all study areas, dead trees larger then 6 inches dbh were counted. The comparison between treated and control sections was significant. Over twice as many trees were dead per hectare in the treated sections as in the control sections. Of the 115 dead trees 103 were Abies concolor, nine were Pinus lambertiana, two were Calocedrus decurrens, and one was Sequoiadendron giganteum. Seven of the P. lambertiana were in the control sections and two trees were in the manipulated section. Both dead C. decurrens trees were in a manipulated section and the giant sequoia was in a manipulated section. Analysis of the 103 dead white fir (Abies concolor) revealed a mortality rate 2.1 times as high in the treated sections as in the control sections (Table 5).
Table 5. Mortality of Abies concolor (6" dbh) in manipulated versus control section, eight to night years after manipulation.
| Area | Number of dead A. concolor per hectare | |
| Manipulated | Control | |
| Trail | 16 | 9.8 |
| Ridge | 18 | 2.5 |
| North | 12 | 7.2 |
| South | 6 | 1.8 |
| Mean | 12.5 | 5.9 |
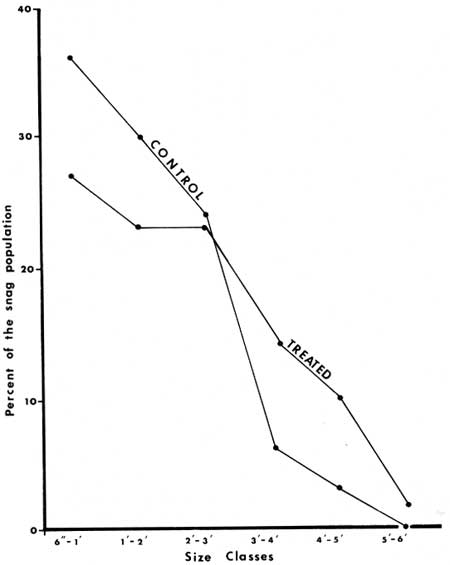
When plotted by size class, the populations of trees showed a difference in the proportion of small versus large individuals (Fig. 34). The treated sections had a greater number of large individuals that had died than the control sections. This was particularly true of the three sections in which heavy equipment had been used. It is possible that mechanical damage occurred, and subsequent invasion by insects or pathogens increased mortality beyond what could be expected from fire alone.
|
| Fig. 34. Mortality in Abies concolor, snags by size class in treated versus control sections. |
Discussion and summary
In light of the patchiness of heat intensities and other irregular factors during surface fires, it is not surprising to find a patchiness in subsequent reproduction of various plant species. Temperatures favorable to seed germination of shrub species, such as Ceanothus spp., will increase their numbers in certain spots, while high temperatures will reduce them. Kilgore and Biswell (1971) reported 16,000 deerbrush (Ceanothus integerrimus) seedlings per hectare on a lightly burned section and only 540 seedlings per hectare on a heavily burned portion, while no seedlings appeared on unburned control plots. The increased number of Ceanothus spp. after fire becomes ecologically important since these nitrogen-fixing shrubs contribute to the chemical nature of the soil (Kilgore 1973).
Two species of the same genus (Galium) appear to have different light requirements and may be adapted by leaf number to their special spectrum of this physical environmental factor. The one that occurs in less light has more leaves.
In a forest successional series, white fir is generally accepted as a late stage tree and tolerant of shade. The present studies quantify this contention and indicate that white fir was most abundant at the lower sunlight values of less than 9% full sunlight, and in fact it was most frequent in plots at only 4% full sunlight. Other species such as Pteridium aquilinum and Rosa californica had high relative frequency and abundance values in plots from 20% to 40% full sunlight, while others such as Viola lobata and Galium sparsiflorum were intermediate.
In addition to tolerating the lower light values of later successional stages, white fir was also quite tolerant of relatively deep duff and litter. On a basis of frequency of occurrence, white fir was present at near expected percentages, whereas several other species, e.g. Adenocaulon bicolor and Hieracium albiflorum, were underrepresented in the thicker litter and duff categories of 6-12 cm in depth, although sample size was insufficient to draw firm conclusions.
There was an increase of annuals in the treated sections of two study areas following the fires or manipulation. After 10 years, however, they were no longer present. This would seem to suggest that ground conditions quickly become unfavorable for germination and/or seedling survival for this type of plant. The fact that none was found where more than 2 cm of litter had accumulated indicates one such condition. Giant sequoia also appear to be a species for which conditions are optimal for only a few years following fire or manipulation (see Chapter 5).
The shrub stage in succession was represented by several species, particularly members of the genera Ceanothus and Ribes. They, along with Phacelia mutabilis, showed a rapid increase after fire and manipulation and then a subsequent decline in numbers. Although they appear to be approaching the population levels pertaining prior to the experimental treatment, it probably will require a few more decades. In a second-growth giant sequoia forest about 1.8 kilometers (1 mi) west of our study area Biswell et al. (1966) suggested that the shrub phase was essentially terminated about 40 to 50 years after logging and fire disturbances in the late 1800s. By 1966 at that site, most shrub species were at less than 5% frequency, while only two shrub species were in the 10% to 20% frequency category (Biswell et al. 1966). In our study areas the relatively rapid passage of annual herbaceous species and of shrub species may be due in large part to the minor alteration in the canopy cover which apparently induced only modest increase in sunlight at the ground level.
In a survey of both control and manipulated sections (with total area of about 11.5 ha) it was discovered that since treatment, twice as many white fir had died in the fire and manipulated sections as in the control sections. In addition there was a disproportionately larger number of large firs that died. More trees in the 1.8 m to 3.6 m (3 ft to 6 ft) classes, which were about 250 years old, died in the manipulated sites than on the non-manipulated. Thus, after about a decade, openings were being created in the crown canopy. This is in contrast to Kilgore's (1973) findings, where only the young firs were affected. Therefore, if another prescribed burn was soon applied to the study areas, the ground conditions might become favorable once again to early stage plants and thus more nearly approximate the space-time mosaic considered by Bonnicksen (1975) as characteristic of this forest ecosystem.
| <<< Previous | <<< Contents >>> | Next >>> |
chap4.htm
Last Updated: 06-Mar-2007