|
NATIONAL PARK SERVICE
Mountain Goats in Olympic National Park: Biology and Management of an Introduced Species |

|
| Mountain Goat Population |
CHAPTER 6:
Habitat Relations, Social Behavior, and Physiological Ecology
D. B. Houston, V. Stevens, and E. G. Schreiner
Habitat Relations
Habitat Utilization
The "psychological bondage" of mountain goats to cliffs and rock outcrops is well known (Brandborg 1955; Geist 1971; Chadwick 1983; Fox 1983; Hutchins and Geist 1987; Fox et al. 1989). Goats are apparently more strongly associated with cliffs than even bighorn sheep (Ovis canadensis; Geist 1971). The ability of goats to occupy and negotiate seemingly impossible terrain in the Olympic Mountains was a wonder to those of us involved in research and management.
Many goats in the Olympic Mountains are seasonally migratory; summer ranges are mostly above 1,520 m and consist of open subalpine and alpine meadows. We know more about habitat utilization by goats during summer than in winter.
Stevens (1979) sampled habitat use by the KR goats from 24 June to 4 September 1978 by recording 5,589 observations of up to 84 individually tagged animals. Bedding (42%) and feeding (40%) composed the activities most observed. Most (55%) of the feeding observations occurred in subalpine meadows (especially Phlox—fescue communities as described by Pfitsch and Bliss 1985; Fig. 27). Observations of feeding (38%) in sparsely vegetated rock-dominated sites (i.e., talus, scree, outcrops—which support Phacelia—Cirsium and Phacelia—Achillea communities) were second. Coniferous forest (7%) was third. South slopes were preferred foraging sites, but goats shifted feeding activity to cooler north slopes during warm days. Goats selected rock-dominated terrain—particularly cliffs—for bedding in 37% of the observations, followed by snowbanks (24%), bare ground (21%), and meadows (14%). Goats favored north slopes for bedding.

|
| Fig. 27. Mountain goats foraging in Avalanche Canyon, Olympic National Park, August 1989. The sparse vegetation is typical of subalpine sites in the dry northeastern Olympic Mountains. Copyright J. E. Burger. |
Intensive use of selected sites led to numerous conspicuous beds and wallows in subalpine and alpine areas. Often it was not possible to distinguish between a bed or wallow at many of these sites. Continued use eliminated vegetation at the site, modified the composition of adjacent vegetation (Reid 1983), and accelerated erosion on particularly steep slopes. Goats maintained site instability by throwing loose soil on themselves while wallowing. Wallowing may aid grooming, may provide thermoregulatory relief (when soils are damp), and may provide relief from biting insects (Brandborg 1955; Stevens 1979). On Klahhane Ridge, goat wallowing occurred throughout summer—often during the warmest time of day (Stevens 1979).
Plots used to describe plant communities and document the extent of herbivory on summer ranges also provided perspective on goat habitat use. Evidence of goat activity (tracks, wallows, droppings) was extensive throughout the 22 areas surveyed in the park and was present in 15 of the 16 community types described, indicating the ability of goats to use a wide variety of plant communities. Pfitsch and Bliss (1985) noted that several herbaceous communities on KR were most heavily grazed when located near cliffs. These observations tended to confirm the strong association of goats with rocks and the seemingly overwhelming influence of these predilections on patterns of vegetation use.
The general locations of winter ranges for four subpopulations were reported from field observations (KR) and from telemetry of 18 radio-instrumented goats (KR, Royal Basin, Constance, Sawtooth Ridge; Stevens 1979). Goats wintered on steep south and southeast slopes below 1,500 m. Radio locations made from fixed-wing aircraft, however, were usually too inaccurate to pinpoint the actual habitat occupied (V. Stevens, personal observation). Fifteen relocations of 11 radio-instrumented goats made by helicopter during winter-spring 1985 in the Mount Appleton area (National Park Service, R. Hoffman, unpublished) showed that 90% of the sites occupied were coniferous forest on steep rocky southeastern or south-facing slopes between 1,200 and 1,500 m. Spring observations from the ground on KR, Baldy Ridge, and Mount Appleton typically showed heavy utilization of virtually all vegetation (including conifers) on cliffs and rock outcrops used during winter; these conditions extended 20-30 m from the cliffs into adjacent forests (D. Houston and R. Hoffman, personal observation; Fig. 28).

|
| Fig. 28. A steep, forested southeast-facing slope that provided winter range for mountain goats in the Cat Creek—Schoeffel Creek area of Olympic National Park. (Photo by R. Hoffman, 2 August 1984) |
The duration of goat occupancy of seasonal ranges varied considerably among years in the Olympic Mountains. Summer ranges were accessible to mountain goats for longer than usual periods during several years in the 1980's because of low fall snowfall, and goats were able to occupy summer ranges earlier in the year because of low snowpacks. On 16 January 1981, we (Houston and V. Stevens) obtained goat composition counts atop Hurricane Ridge under near-summerlike conditions. During typical winters, snow would have displaced goats from these high-elevation meadows. Crude indices to this variability are represented by the coefficients of variation for the January (46.7%, 1960-92) and April (51.8%, 1958-92) snow-depth measurements at Hurricane Ridge. Snow depths ranged from 18 to 234 cm in January and from 0 to 277 cm in April.
Studies of habitat selection by goats in coastal areas of British Columbia and southeastern Alaska have shown extensive winter use of coniferous forest on and adjacent to steep south slopes and cliffs (Hebert and Turnbull 1977; Schoen and Kirchoff 1981; Fox 1983; Smith 1984, 1986; Fox et al. 1989). Overstory trees in mature coniferous forests intercept snow and provide foraging sites that would be unavailable otherwise (National Council of the Paper Industry for Air and Stream Improvement 1989). Our limited observations suggest that goats use forests on winter ranges throughout the Olympic Mountains.
Goat Diets
Seasonal Diets
Seasonal diets of the KR subpopulation were examined to aid interpretation of the effects of goats on vegetation. Microhistological examination of fecal material was used to identify plant taxa in the diets (Vavra and Holechek 1980; Leslie et al. 1983).9
9Analyses were performed at the Eastern Oregon Agricultural Research Station, Union, Oregon. Composite samples (5 g from each pellet group) were made for six seasonal periods from a sample of 245 pellet groups collected October 1977-April 1982 (January-March, 29 pellet groups; April-May, 25; June, 41; July, 39; August-September, 61; October-December, 50). Samples from fall 1977 to winter 1981 were collected by V. Stevens.
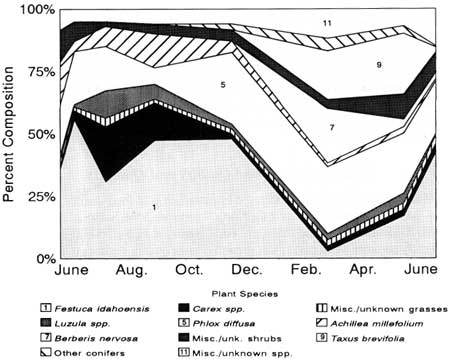
Of the nearly 200 plant species known on KR, 39 taxa were identified in seasonal diets (Table 19). Seven taxa made up at least 5% of the diets for one or more seasonal periods (Fig. 29).
Table 19. Plant taxa identified in mountain goat (Oreamnos americanus) diets from Klahhane Ridge.a
| Forage class | Scientific name | Common name |
| Grasses and grasslike plants | ||
| Agrostis variabilis | Mountain bentgrass | |
| Bromus sitchensis | Alaska brome | |
| Carex spp. | Sedges | |
| Dactylis glomerata | Orchardgrass | |
| Elymus glaucusb | Blue wildrye | |
| Festuca idahoensis | Idaho fescue | |
| Festuca ovina | Sheep fescue | |
| Juncus spp. | Rushes | |
| Luzula spp. | Woodrush | |
| Phleum alpinumb | Alpine timothy | |
| Poa spp. | Bluegrasses | |
| Sitanion hystrix | Squirreltail | |
| Stipa occidentalis | Western needlegrass | |
| Forbs | ||
| Achillea millefolium | Yarrow | |
| Arabis spp. | Rockcress | |
| Campanula piperi | Piper's bellflower | |
| Cheilanthes gracillimab | Lace lip-fern | |
| Phlox diffusa | Spreading phlox | |
| Polystichum munitum | Swordfern | |
| Pteridium aquilinumb | Bracken | |
| Rubus spp. | Brambles | |
| Browse | ||
| Acer circinatumb | Vine maple | |
| Alnus rubrab | Red alder | |
| Arbutus menziesii | Pacific madrone | |
| Berberis nervosa | Oregon grape | |
| Douglasia laevigata | Smooth douglasia | |
| Gaultheria shallonb | Salal | |
| Pachistitna myrsinites | Oregon boxwood | |
| Symphoricarpos mollisb | Snowberry | |
| Vaccinium membranaceumb | Thin-leafed huckleberry | |
| Vaccinium parvifolium | Red huckleberry | |
| Conifers | ||
| Abies lasiocarpa | Subalpine fir | |
| Pinus monticola | Western white pine | |
| Pseudotsuga menziesii | Douglas-fir | |
| Taxus brevifolia | Pacific yew | |
| Thuja plicata | Western redcedar | |
| Tsuga heterophylla | Western hemlock | |
| Tsuga mertensiana | Mountain hemlock | |
| Miscellaneous | ||
| Bryophytes | Mosses | |
aTaxa identified in 245 fecal pellet samples from goats collected during late winter or spring unless otherwise noted. bTaxa identified in 9 rumen samples from goats collected during late winter or spring. | ||
|
| Fig. 29. Seasonal diets of mountain goats from Klahhane Ridge as determined from analysis of 245 pellet group samples. Plant species making up at least 5% of the diets during one or more seasonal periods are identified. |
Seasonal changes of forage classes in the diet were similar for goats elsewhere and for other medium-sized ungulates in mountainous areas of the North Temperate Zone. Grasses and grasslike plants composed about 60% of June-November diets; three forb species contributed much of the remainder. Shrubs and conifers predominated during winter. At the species level, Phlox diffusa, Taxus brevifolia, Berberis nervosa, and miscellaneous shrubs (mainly Pachistima myrsinites and Vaccinium spp.) were important components of the winter diet. Festuca idahoensis and Carex spp. dominated summer-fall diets. Miscellaneous species included small amounts of six forbs in summer and Polystichum munitum (4.5%) and mosses (2.4%) in winter.
Only one of eight endemic plant taxa known on KR was recorded in the mountain goat diet. Campanula piperi made up 0.4% of the diet from January to March and 0.6% from August to September. Three other endemics—Senecio neowebsteri, Astragalus australis var. olympicus, and Aster paucicapitatus—were known to be grazed by goats (see Chapter 12; Pike 1981). Thus, the endemic plants were so uncommon that they seemed to represent only incidental items in the diet of mountain goats.
We add several caveats to the interpretation of data on mountain goat diets. Although samples were corrected for differential digestibility of species (Leslie et al. 1983), microhistological analyses still provided only approximate diet composition. For example, forbs are highly digestible and consequently are often underestimated (Vavra et al. 1978; Samuel and Howard 1983). Field observations and indices of summer grazing on the KR vegetation plots (Pfitsch and Bliss 1985) showed widespread use by goats of many forbs that were present at low densities, and yet few of these were detected by fecal analyses. Parkwide, we expect great variation in the plant species consumed by goats—Festuca idahoensis, for example, is uncommon on ranges in the interior of the park. Considerations of the vegetation parkwide suggested, however, that the relative importance of forage classes would be general, even though the particular species in diets differed among subpopulations.
Finally, samples used in this analysis were collected during unusually mild winters when precipitation was below normal. High-elevation meadows were unusually available to wintering goats. In years with normal to heavy snow accumulations, diets dominated by the shrubs and conifers characteristic of January to March would probably also typify the October to December period.
The plant community survey plots provided additional insight into summer goat diets parkwide. Many different plant species were consumed in areas of high goat density,10 although the particular species selected differed markedly among sites (see Chapter 9). These observations provided additional confirmation that goats in the Olympic Mountains were generalist herbivores and indicated that requirements for rocky habitats apparently overrode needs for particular forage species.
10Plants were assumed to be eaten mainly by goats but included grazing by other mammalian herbivores as well.
Seasonality of Food Quality and Quantity
Marked seasonal variation occurred in mountain goat food quality and quantity—parallel to most ungulates in the North Temperate Zone. Seasonal differences are well documented in the literature. Food quality is highest during late spring and early summer when rapid plant growth occurs; quality declines over summer and fall as vegetation matures; quality is lowest during winter when plants are dormant. Changes in food quantity generally lag behind food quality. Quantity is greatest in middle to late summer. Quantity is most limited during winter when goat distribution is constrained by snow accumulation and the animals feed on a fixed, less diverse, and declining resource base of dormant plants.
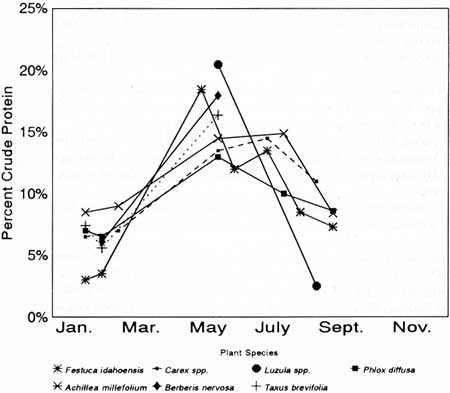
Definitions of food quality vary, but the concentration of nitrogen, reported as percent crude protein, is a useful index for ungulate foods if used cautiously (Crampton and Harris 1969; Cowan et al. 1970; Sinclair 1977). Seasonal crude protein concentration of seven plant species important in diets of KR goats showed that highest levels (around 15-20%) usually occurred during May-June and lowest levels (3-7%) during winter (Fig. 30).
|
| Fig. 30. Crude protein levels in seasonally important foods of mountain goats on Klahhane Ridge. Crude protein calculated as percent nitrogen x 6.25. |
Plants were assumed to be eaten mainly by goats but included grazing by other mammalian herbivores as well.
Seasonal changes in the quality of goat diets, as indicated by crude protein values in forage, also were reflected in fecal nitrogen (N) levels. Fecal N averaged 1.69 ± 0.26% on KR during October to March 1977-81 and was 2.29 ± 0.41% for June samples (Stevens 1983).11 N values from goats on three other summer ranges in the park (Mount Constance, Mount Appleton, and Royal Basin) averaged 3.27 ± 0.22% during 1980 and were greater than values from KR (Stevens 1983). Data on demography and goat condition led Stevens (1983) to conclude that these three subpopulations were less intensely resource-limited than goats on KR, and this was reflected in the fecal N values.
11There is controversy over the validity of fecal N as an index of diet quality because tannins in some dicotyledonous forages may reduce protein digestibility (Hobbs 1987; Leslie and Starkey 1987; Robbins et al. 1987; Irwin et at. 1993). Here, values are used to reflect only broad seasonal differences in diet.
Measures of available food quantity are limited. Standing crop for nine major plant communities on 125 ha of summer range above 1,400 m on KR was calculated at 154,610 kg (~1,237 kg/ha) during 1979 (Pfitsch and Bliss 1985). Goats consumed a small proportion of the standing crop.
Diets Elsewhere
About 20 studies reported diets of mountain goats over the past 40 years. Differences in design and sampling methods, however, limit comparisons. Information on diets has been derived from rumen samples, range surveys that attempted to relate plant abundance to goat utilization, timed observations of feeding animals, examinations of feeding sites, and more recently, microhistological examination of fecal samples. All techniques are prone to sampling errors and biases (e.g., Nelson and Leege 1982).
Even with the limitations, several generalizations on food habits are useful:
1. Over their geographic range, goats consume a remarkable variety of plant species, including mosses, lichens, ferns, grasses, sedges, rushes, forbs, and a host of coniferous, deciduous, and evergreen trees and shrubs;
2. Spring and summer diets are usually dominated by grasses or forbs;
3. Winter diets are frequently dominated by browse species, including conifers;
4. The proportions of browse in diets increase sharply during severe winters; and
5. Conifers are often important components of winter diets, even during mild conditions when alternate forages are seemingly available.
These interpretations are drawn from our studies and from Casebeer et al. (1950), Brandborg (1955), Saunders (1955), Hibbs (1967), Geist (1971), Hjeljord (1973), Kuck (1973), Johnson et al. (1978), Thompson (1980), Adams and Bailey (1983), Campbell and Johnson (1983), Smith (1986), Fox and Smith (1988), Fox et al. (1989), and Cowan (I. M. Cowan, Wildlife Service, Department of Mines and Resources, Ottawa, Canada, unpublished report, 1944).
Mountain goats have anatomical specializations usually associated with ungulates that forage selectively—they possess relatively narrow muzzles and incisors (Janis and Ehrhardt 1988). Thus, in addition to their ability to consume many species, foraging goats frequently select specific, highly nutritious plant parts—particularly flowers, seed heads, and growing leaves or leaf blades (Saunders 1955; Hjeljord 1973; Olmsted 1979; Thompson 1980; Dailey et al. 1984). Moreover, wintering goats dig up and consume energy-rich, belowground plant structures, including fern rhizomes, roots of perennial forbs, bulbs, and tubers (Hjeljord 1973; Chadwick 1983; D. B. Houston, personal observation).
Social Behavior
Mountain goat society is matrifocal, with groups of females, kids, and young males generally living apart from groups of adult males, except during rut (Geist 1964; DeBock 1970; Chadwick 1977). Rut occurs from late November to early December, with large, dominant males doing much of the breeding (Brandborg 1955; Geist 1964; Hoffman 1987; Chadwick 1973). Male-male dominance is established by threat displays; fighting is apparently uncommon but can be deadly (Geist 1967; DeBock 1970; Dane 1977). Males tend individual females, as tending pairs, during the latter's 48-h period of sexual receptivity (Hutchins et al. 1987). Recurrent estrus may follow at intervals of about 20 days. Gestation is about 186 days, and females give birth in late May. Females remain in seclusion for up to 17 days postpartum as maternal-young bonds develop (Hutchins et al. 1987). Neonates are precocial followers (as opposed to hiders; Lent 1974) capable of negotiating difficult terrain after just a few days. Goat kids, as followers, grow initially more slowly than mule deer fawns (hiders; Carl and Robbins 1988). Kids nurse frequently but consume relatively less milk per nursing bout than do deer fawns.
Social Organization and Group Size
Goat society is ordered along dominance hierarchies, with dominant individuals frequently being older and larger (DeBock 1970; Chadwick 1977; Risenhoover and Bailey 1985). In the Olympic Mountains and in several other locations, large adult males seem to be dominant over females (Rideout 1974; Dane 1977; Stevens 1983); however, the extent and circumstances of male dominance may vary among populations (Geist 1964; De Bock 1970; Chadwick 1977, 1983; Masteller and Bailey 1988). Overt aggressive behavior toward social subordinates increases in winter when resources are limited (Petocz 1973). Conversely, agonistic behavior between goats apparently declines with increasing group size in summer when resources are abundant (Risenhoover and Bailey 1985).
Females that do not give birth may retain their yearling or even 2-year-old offspring at heel. Those with retained offspring, plus females with kids of the year, represent the nuclei of nursery groups (Hutchins 1984). Although associations among adults in nursery groups were not measured quantitatively in Olympic National Park, group constancy seemed to be low (Stevens and Driver 1978; M. Hutchins, American Zoo and Aquarium Association, Bethesda, Maryland, personal communication, 1988). In Glacier National Park, associations between tagged female goats also were low (Singer and Doherty 1985).
Mean group size for goats on summer ranges in the Olympic Mountains was 2.74 ± 3.51 SD (range 1-32, n = 162 groups) in July 1983 and 2.05 ± 1.80 (range 1-12, n = 84 groups) in 1990 and did not differ between years (Mann-Whitney, P > 0.05). In 1983, 42 nursery groups (i.e., those containing at least one female with kid-at-heel) were larger and averaged 5.5 ± 5.46 goats; in 1990, 25 nursery groups averaged 3.8 ± 2.42 goats.
We recorded goat density and group sizes on KR during June censuses from 1981 to 1986 (Table 20). Mean group size was positively associated with June population size (r2 = 0.88, P < 0.05) and declined precipitously during the goat reductions. Nursery groups showed similar changes; the largest such group recorded each year for 1981-86 was 31, 24, 33, 7, 6, and 6. The larger nursery groups (20 or more) often occurred in open meadows some distance from escape terrain. Risenhoover and Bailey (1985) suggested that large groups permit more efficient foraging in open terrain because individual females could devote more time to foraging and less time to alert behavior.
Table 20. Mountain goat (Oreamnos americanus) density and group size, Klahhane Ridge, 1981-1986.a
| Group size | ||||
| Year | Goats/ha | n |  |
CVb |
| 1981 | 0.077 | 35 | 4.40 | 130 |
| 1982 | 0.063 | 29 | 4.28 | 107 |
| 1983 | 0.045 | 22 | 4.09 | 167 |
| 1984 | 0.024 | 27 | 1.85 | 5 |
| 1985 | 0.012 | 16 | 1.40 | 5 |
| 1986 | 0.014 | 14 | 2.10 | 77 |
aObserved during June census flights that surveyed 1,997 ha. bCV = coefficient of variation. | ||||
Limited November-to-March observations on KR suggested that winter groups were at least as small as in summer. Mean winter group size was 1.8 ± 2.21 goats (range 1-7, n = 31 groups).
Land Tenure and Home Range
Mountain goats in the Olympic Mountains showed fidelity to well established seasonal home ranges. One hundred twenty-five goats tagged on KR were reobserved for 2 or more years following capture. About 84% showed fidelity to summer ranges—several for 5-10 years (calculated from Stevens [1980, 1983] and subsequent National Park Service observations). Fifty-five of 57 tagged goats (97%) in 8 other subpopulations showed fidelity to summer ranges. Levels of fidelity to winter ranges were unknown but were probably similarly high because habitat was more limited and resources were in short supply. Goats elsewhere showed fidelity to summer (Rideout 1977; Thompson 1980) and winter ranges (Smith 1976; Kuck 1977; Schoen and Kirchoff 1981; Smith and Raedeke 1982); fidelity to winter ranges seemed more pronounced among females than among adult males. Indeed, females in some populations exhibited facultative territoriality in winter and seemed to lay exclusive claim to the highest quality cliff habitats (Hutchins and Geist 1987).
Size of seasonal or year-round home ranges of goats was not calculated during our studies. Radio-instrumented goats in other populations had small annual home ranges of 6-25 km2 (Rideout 1977; Thompson 1980; Singer and Doherty 1985). Winter home ranges were much reduced—in some instances to 1 km2 (Smith 1976).
Physiological Ecology
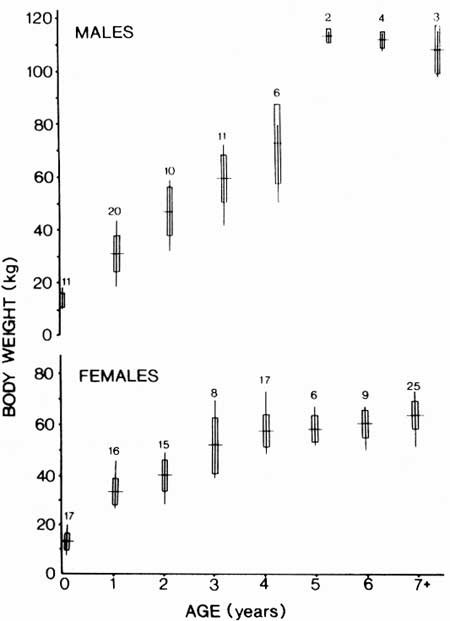
We contrasted live weights of wild and captive-reared mountain goats from KR to determine if weight differences might reflect the extent of resource limitation (Houston et al. 1989). Age-specific body weights of 180 wild goats varied considerably; coefficients of variation ranged from 15 to 22% of the mean body mass for the 1- to 3-year-old males and females (Fig. 31). Goat population size contributed to this variation because mass increased as goat density was reduced (see Chapter 5). Wild females reached maximum body mass at 3 or 4 years; wild males at 5 years. Mass of wild males and females did not differ from kid through 3-year-old classes, but pronounced sexual dimorphism in body mass occurred by age 4. Body mass of wild yearlings, 2-year-olds, and 3-year-olds in July was only 55-68% of that of captive-reared animals of the same age (Houston et al. 1989).
|
| Fig. 31. Body weights of male and female mountain goats in relation to age, Klahhane Ridge. Values represent sample size with mean, range, and 1 SD. |
We know little about the seasonal patterns of growth in wild goats. Captive goats, however, gain weight rapidly from May to October, and either remain stable or lose mass from November to April (Houston et al. 1989). Yearling and older males lost weight beginning in late October (about 1 month before rut), and the loss became more pronounced with age. Captive males averaged 75 kg and females averaged 70 kg before breeding at about 1.5 years of age. Captive males reached approximately 65% of their mean adult body mass by their first breeding season; captive females, 80%.
Even when fed high quality food year-round, growth of captive goats varied seasonally (Houston et al. 1989), a pattern common to other ungulates living in the North Temperate Zone (e.g., Wood et al. 1962). Wild goats on KR required considerably longer than did captives to reach the same body mass. Sexual maturity occurred earlier in captive as compared to wild goats.
Little is known about the nutritional physiology of mountain goats. Theory and studies of other species suggest that medium to small ungulates, such as mountain goats, should select high quality foods that are digested rapidly (e.g., Bell 1971; McNaughton and Georgiadis 1986; Baker and Hobbs 1987). Such selection is necessary to maintain the relatively high metabolic rates of the smaller species. Limited dietary studies suggest that goats might follow this strategy, but no experimental studies of digestive efficiency are available.
Mineral requirements are poorly understood. Mountain goats show much lower dietary selenium requirements than domestic animals, and this has been interpreted as an adaptation to low-selenium environments (Robbins et al. 1985). Sodium may be deficient in soils and vegetation of alpine areas that receive heavy precipitation, and salt craving displayed by goats in the Olympic Mountains in spring and early summer may reflect reduced availability of this element (Robbins 1983).
Interpretation
The remarkably catholic diet and versatile foraging behavior of the mountain goat seem to compensate for their comparatively narrow habitat preference (Geist 1962). The ability of the mountain goat to consume a wide variety of plants has surely contributed to successful colonization of the ecologically diverse Olympic Mountains in less than 60 years. Slower body growth and associated lag in sexual maturity exhibited by wild goats, compared with captive-reared animals, underscores our contention that at least some subpopulations in the Olympics were resource-limited.
Geist (1987) argues persuasively that the duration of the annual productivity pulse of vegetation (i.e., the period of release from shortages in quality and quantity of food resources) has been a powerful force in shaping the ecology and evolution of Pleistocene herbivores. In the mountain goat, life habits, physiology (including seasonality of growth), and behavior seem closely aligned with this seasonal pulse. In the Olympic Mountains, goat migrations to resource-rich—but only seasonally available—summer ranges precede periods of growth, lactation, and fat accumulation. Migrations prolong access to abundant, high quality resources by effectively extending the productivity pulse.
The ability to persist in small groups organized along dominance hierarchies permits mountain goats to successfully exploit isolated, small pockets of resources. Among social animals in general, dominant females often show superior breeding success (Clutton-Brock and Albon 1985). This may be true for mountain goats, but it has not been demonstrated.
| <<< Previous | <<< Contents >>> | Next >>> |
chap6.htm
Last Updated: 12-Dec-2007