|
HAWAI`I VOLCANOES
Invasion and Recovery of Vegetation after a Volcanic Eruption in Hawaii NPS Scientific Monograph No. 5 |

|
CHAPTER 5:
Plant Invasion Patterns (continued)
Spatter Area with Tree Snags (Habitat 3)
This is the relatively small, narrow, and elongated habitat adjacent east of the cinder cone (habitat 2) and adjacent south of Kilauea Iki (Fig. 2). It was defined in outline by the presence of Metrosideros tree snags that remained standing or fell down after the flowing-hot pyroclastic fallout was deposited. The new surface consisted of "welded spatter," i.e., relatively large chunks of cinder-like particles that were still soft from heat and became welded upon deposition. The presence of completely defoliated trees (=snags) provided a convenient, mapable outline of habitat 3, which was bordered on the east side by the surviving, still well-foliated Metrosideros stand. On the north side it was bordered by the lower slope of the cinder cone habitat. In the first year after the eruption, the National Park Service built a board walk through habitat 3 for the use of visitors.
Vegetation was sampled by two transects: BB', 60 m long, with six contiguous 10 x 10-m plots running east-west from the intact forest to the cinder cone slope, and EE', with 15 contiguous 10 x 10-m plots running north-south on the cinder cone side, 5-10 m parallel to the board walk.
Progress of invasion
A directional advance of invasion of new species across habitat 3 was recorded by species presence in the six 10 x 10-m-contiguous quadrats running from the undisturbed forest edge to the cinder cone slope, over a distance of 60 m (Table 3).
It can be seen that algae were present over the 60-m belt-transect in all six quadrats beginning with the first year after the eruption.
Mosses arrived in the second year within the first 20 m off the forest edge and then spread across habitat 3 by advancing less than 10 m per year. In year 7, mosses were found in all six quadrats.
The lichen Stereocaulon volcani arrived together with the mosses in the second year and spread faster than 10 m per year across habitat 3.
The fern Nephrolepis exaltata arrived in the first year and then spread with a rate similar to Stereocaulon across the habitat.
Because of the high number of seed plant species that arrived early in the habitat, this group has been further separated into grasses, sedges, forbs, shrubs, and tree seedlings in Table 3.
Grasses arrived in the first year and spread with much the same speed (i.e., 10 m/year) across habitat 3. The first grasses were three exotic perennials: Setaria geniculata, Paspalum dilatatum, and Sporobolus africanus.
TABLE 3. Progression of invaders from undisturbed forest edge across habitat 3 towards cinder cone along the 60-m belt-transect BB' (Fig. 2).
| Life forms | Year after eruption |
1 1960 | 2 1961 | 3 1962 |
4 1963 | 7 1966 | 9 1968 |
| Algae | 60m | 60m | 60m | 60m | 60m | 60m | |
| Mosses | — | 20 m | 20 m | 30 m | 60 m | 60 m | |
| Lichens | — | 30m | 30m | 50m | 60m | 60m | |
| Ferns | 20m | 20m | 20m | 50m | 60m | 60m | |
| Grasses | 10m | 20m | 30m | 40m | 50m | 60m | |
| Sedges | |||||||
| native | — | — | — | — | — | 50 m | |
| exotic | — | — | 20 m | 20 m | 50 m | 50 m | |
| Forbs | |||||||
| native | 10m | 10m | 20m | 20m | 20m | 50m | |
| exotic | 10m | 20m | 30m | 60m | 60m | 60m | |
| Shrubs | |||||||
| native | — | 20 m | 30 m | 50 m | 60 m | 60 m | |
| exotic | 10m | 20m | 30m | 40m | 30m | 40m | |
| Tree seedlings | |||||||
| native | — | — | 20m | 30m | 60m | 60m | |
| exotic | — | — | 20 m | 40 m | 60 m | 60 m | |
A separation into native and exotic species aids in the interpretation of the result of the following seed plant groups:
Exotic sedges arrived after the second year and penetrated 50 m into habitat 3 in the seventh year, where they remained through the ninth year. At that time, a native sedge, Carex wahuensis, made its entrance at the 50-m part of the transect, but this species occurred only in one of the five quadrats.
Forbs occurred together with the algae, ferns, and grasses in the first year after the eruption. One of these was a native species, Astelia menziesiana; another, an exotic, Eupatorium riparium. Exotic herbs penetrated rapidly across the transect, reaching the 60-m part in the fourth year after the eruption. Astelia advanced another 10 m into habitat 3 in the third year. It remained in the first two quadrats throughout the 9-year observational period. But a second native herb, Hedyotis centranthoides, was recorded in two quadrats in the ninth year. One of these was the fifth quadrat.
An exotic shrub, Rubus rosaefolius, appeared in the first year after the eruption in the first quadrat. Thereafter, it was joined by several other species, for example, Rubus penetrans, in the second year. However, this group did not advance beyond the 40-m section of the transect.
In contrast, a native shrub, Dubautia scabra, did not appear until the second year. It was then joined by other native shrubs in the fourth year, notably Vaccinium reticulatum and V. calycinum. Both Dubautia and V. reticulatum spread rapidly to the sixth plot, where they were recorded in the seventh year.
Tree seedlings did not arrive until the third year. These included only two species, the native Metrosideros polymorpha and the introduced weed-tree Buddleja asiatica. Both trees spread with the same rate across the 60-m transect and were found in the sixth plot in the seventh year (1966).
Although directional progress was not measured along transect EE' (Fig. 2), plants were observed to arrive slowly in the northeast sector and to progress toward the southwest.
Floristic pattern in comparison to habitats 1 and 2
As pointed out previously, the arrival sequence of life forms and species differed from those in habitats 1 and 2. Appendix III shows that seed plants were among the first invaders, together with the alga Stigonema panniforme and the fern Nephrolepis exaltata. Mosses and lichens arrived in the second year. Thus, within the group of cryptogams the sequence of arrival remained similar to that on the two previously discussed habitats, but the arrival of seed plants together with the cryptogams is a significant departure for habitat 3. It is also of interest that these first seed plant pioneers were all exotics except one, the forb Astelia menziesiana. This species grows normally as an epiphyte on very old Metrosideros trees in the montane rain forest. In habitat 3, Astelia grew directly on the ash. The exotics that appeared as first arrivals in year 1 were a shrub (Rubus rosaefolius), three grasses (Paspalum dilatatum, Setaria geniculata, and Sporobolus africanus), and one forb (Eupatorium riparium). Except for the shrub life form, there are very few native species among the grasses and forbs in this climatic zone (Doty and Mueller-Dombois 1966) that could be expected as early invaders.
A second, remarkable departure from the species invasion pattern of habitats 1 and 2 is the very rapid increase in the number of species in habitat 3. They increased from 8 in year 1 to 64 in year 9. This increase in species is particularly noteworthy in the group of mosses, grasses, sedges, and forbs.
The first three moss species in Appendix III are the common pioneer mosses found also in habitats 1 and 2; the other 12 mosses are unique to this habitat. The increase in number of moss species was explosive in year 7. This correlates with a rather sudden recovery of the woody plants that began to provide shade over much of the spatter surface on habitat 3. This woody-plant recovery is documented by the photographic record (Fig. 9). Except for Ceratodon purpureus, the mosses that invaded habitat 3 in year 7 and 9 are species of the rain forest. Thus, the sudden increase in moss species indicates a significant change in microclimate near the ground in habitat 3. The change to a continually more moist microclimate near the ground began in year 4 with the arrival of Bryum crassicostatum and Macromitrium owaihense.
The second major floristic difference from the first two habitats is in the early establishment of a species-rich herbaceous component in habitat 3. This also indicates much more mesic conditions. Only two species in the herbaceous component are endemic, the sedge Carex wahuensis and the forb Astelia menziesiana. All 9 grasses, 3 sedges, and 20 forb species are exotics. It seems apparent that these exotic herbs fill an almost empty niche in the invasion process on new volcanic materials. Some of these had already made their entrance on the crater floor (habitat 1), and it is to be expected that many, if not all, of the exotic herbs listed in Appendix III will also become established in the other new habitats in the rain-forest climate.
Succession
The species frequency data in Appendix III shows a rapid rate of spread over habitat 3 for a number of species. For example, the algae were found in all 21 quadrats in year 4; the moss Campylopus exasperatus and the lichen Cladonia skottsbergii, in year 7; and the pioneer fern Nephrolepis exaltata had spread across nearly the entire habitat in year 9. This indicates a much faster rate of occupation than was shown for the cinder cone (habitat 2).
A few species showed a decline in territorial gains over the 9-year period in habitat 3. This is significant because the decline may indicate the beginnings of a succession.
A definite territorial reduction was shown by the moss Campylopus densifolius, which showed its peak spread with 95% frequency in year 7 and thereafter was found only in 10% of the plots. Here, a successional replacement with one or several of the rain-forest mosses is indicated in the ground layer.
Among the shrubs, a definite territorial decline was shown in the two Rubus species (R. rosaefolius and R. penetrans) from their peak distribution in year 4. It was observed that spaces occupied by certain individuals of these exotic species were later invaded by individuals of the native shrubs Dubautia scabra and Vaccinium reticulatum. Thus, in these cases, a successional replacement of exotic by native shrubs was indicated. The same phenomenon was observed with the exotic weed-tree Buddleja asiatica, which showed significant territorial reduction in year 9. A large number of mature individuals of Buddleja were dying off rather suddenly. The Buddleja-dying took place particularly among those individuals that grew at the bases of Metrosideros snags that had resprouted from the base. The possibility of competitive replacement of Buddleja trees by Metrosideros trees was indicated.
Among the grasses, definite territorial reductions occurred in Paspalum dilatatum and Setaria geniculata. Sporobolus africanus disappeared completely from the plots in year 7. These three grass species were those that occurred in year 1. Their peak distribution was in year 4. Their decline coincides with the appearance of the mosses, which indicated a change in microclimate near the ground caused by increased shading of the woody plants. However, the decline of the grasses was noted also in unshaded places. Here the grasses Holcus lanatus and Pennisetum clandestinum and the sedge Cyperus polystachyos took over some of the space of the above declining species, indicating competitive replacement.
Similar replacement relations were observed among the forbs. After year 4, the first invader Eupatorium riparium lost much of its territory by die-back. Significant decreases occurred also in Geranium carolinianum. Sonchus oleraceus, and Erechtites valerianaefolia. Later invaders and definite increasers were Commelina diffusa, Hypochoeris radicata, and Epilobium cinereum.
Therefore, succession through competition was definitely indicated in habitat 3. While among the woody plants there was a trend toward greater habitat occupation by native plants at the expense of exotic shrubs, the competition in the herbaceous group was strictly among exotics.
Pumice Area with Tree Snags (Habitat 4)
This habitat consists of the area where the first snag tops of buried trees began to appear above the pumice blanket. The boundary extends to the survival tree line of habitat 5, where the maximum pumice depth was about 3 m (Figs. 2 and 3). A single tree fern (Cibotium glaucum) was the only plant that was observed to survive the deep burial and pumice blast. In the ninth year this plant still displayed good vigor, but no additional specimens had appeared. Since most of this habitat area already lies in the area of the former seasonal forest (Fig. 2), no additional tree fern specimens were expected. The tree fern C. glaucum is a characteristic plant of the rain forest.
Arrival pattern in comparison to habitats 1, 2, and 3
The general direction of invasion was from both the eastern and western boundaries of the habitat toward the center, but the new plants became established in a widely scattered formation.
Appendix IV shows the invading species for the 72 quadrats. No new plant was recorded in year 1. In year 2, seed plants became established together with the first cryptogams. Simultaneous arrival of seed plants and cryptogams was also noted in habitat 3. Both these habitats (3 and 4) were characterized by the standing tree snags. Therefore, there is a correlation of immediate seed plant establishment and presence of significant organic residues of a former vegetation (here, the tree snags) on new volcanic surfaces. The pioneer seed plants arriving in year 2 in habitat 4 were the exotic shrub Rubus rosaefolius, the native tree Metrosideros polymorpha (as seedling), and the composite weed Erechtites valerianaefolia.
The cryptogam-arrival sequence was similar to that on the previously discussed habitats, except that the potential semi-tree fern Sadleria cyatheoides appeared together with Nephrolepis exaltata in year 2. In habitats 1 and 2, Sadleria appeared a year after Nephrolepis. Again, the lichen Stereocaulon volcani arrived a year after the other cryptogams (algae, mosses, and ferns).
Among the seed plants, several more woody species became established in year 3 (two exotics Rubus penetrans and Buddleja asiatica and one native Dubautia scabra), while only two additional forbs appeared. These were also weedy composites (Sonchus oleraceus and Hypochoeris radicata). In contrast to habitat 3, grasses arrived very late, only in year 7 (Andropogon virginicus). Only one more grass species (Holcus lanatus) became associated in year 9. Also, the rain-forest mosses found in habitat 3 never arrived in habitat 4.
The comparison with habitat 3 is of interest because both habitats had the tree snags in common. Table 4 shows the number of seed plant species found in the two habitats.
Habitat 3 received more rainfall than habitat 4 (see rainfall gradient, Fig. 3). Habitat 3 was in the rain-forest terrain and habitat 4 in what was formerly mapped as seasonal forest (see boundary in Fig. 2). This difference in moisture may in part explain the different pattern in seed plant establishment on the two habitats. Another difference is in the greater water-holding capacity of the spatter in habitat 3 as compared to that of the pumice in habitat 4. This will be discussed later. The greater distance away from an intact, surviving stand, of course, also was important. The disseminules reaching habitat 4 had to be carried over a greater distance. This may have eliminated the establishment of a number of grass species in habitat 4 that were present in habitat 3. However, the disseminule dispersal limitation was probably a lesser factor among the moss species. The absence of rain-forest mosses in habitat 4 points to water rather than to dispersal as the more important limiting factor in early species diversity.
TABLE 4. Number of seed plant species in the two pyroclastic habitats with tree snags (habitats 3 and 4).
| Life form and habitat | Year | 1 1960 | 2 1961 | 3 1962 |
4 1963 | 7 1966 | 9 1968 |
| Woody plants | |||||||
| habitat 3 | 1 | 3 | 6 | 9 | 9 | 11 | |
| habitat 4 | — | 2 | 5 | 5 | 5 | 7 | |
| Grasses | |||||||
| habitat 3 | 3 | 3 | 5 | 8 | 6 | 7 | |
| habitat 4 | — | — | — | — | 1 | 2 | |
| Sedges | |||||||
| habitat 3 | — | — | 2 | 3 | 3 | 4 | |
| habitat 4 | — | — | — | — | — | — | |
| Forbs | |||||||
| habitat 3 | 2 | 4 | 4 | 14 | 18 | 21 | |
| habitat 4 | — | 1 | 3 | 5 | 9 | 12 | |
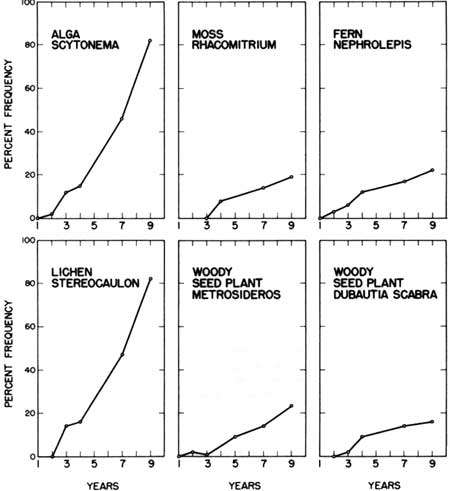
Quantitative spread of species in comparison to habitat 2
Figure 7 shows the rates of spread in 10 x 10 m squares of six selected species over the surface area of habitat 4. The six species were the most widely distributed in habitat 4. A comparison of Fig. 7 with Fig. 6 shows that the rates of spread were quite similar on both the cinder cone and pumice-with-tree-snags habitat. The pioneer algae and lichens on both habitats showed the fastest rate of spread. The pioneer moss Rhacomitrium and the woody plant seedlings also displayed similar, but much slower, rates than the algae and lichens. The fern Nephrolepis exaltata progressed only half as rapidly in habitat 4 (Fig. 7) as on the cinder cone (Fig. 6). This is undoubtedly related to the number of fissures in the substrate surface. There were many more cracks and crevasses on the cinder cone surface than on the pumice-with-snags habitat. While Nephrolepis became established also on the level pumice surface, it takes a foothold much more readily in cracks, as noted particularly for habitat 1 (Fig. 5).
|
| Fig. 7. Rate of spread across pumice-with-snags habitat (4) for selected species in five life-form groups. |
Pumice Area with Surviving Trees (Habitat 5)
This habitat was covered with a blanket of pumice varying from 3 m depth at its boundary with habitat 4 to about 25 cm depth at its boundary with habitat 6 (Figs. 2 and 3).
Under this ash deposit, most Metrosideros trees plus 22 other species survived. Their recovery will be discussed later.
A separate record was kept of new invaders and survival plants. The list of new invaders is shown in Appendix V.
Similarly, as in habitat 4, no new invader was noted in year 1. In year 2, the typical pioneer cryptogams arrived (an alga, Stigonema; a moss, Campylopus densifolius; and the fern, Nephrolepis exaltata). Again, the first lichen (Stereocaulon volcani) was the last of the cryptogamic life forms, arriving in year 3, but it was noted all over the habitat. Of significance also is the arrival of seed plants together with the pioneer cryptogams in the first arrival year. Thus, the same life-form arrival sequence was recorded as in the habitats with snags (3 and 4). The first seed plants were an exotic sedge (Cyperus brevifolius) and an exotic forb (Eupatorium riparium). Eupatorium riparium was also the first forb to vigorously invade habitat 3, and C. brevifolius also was the first sedge together with C. rotundus to invade habitat 3 in year 3.
There are other floristic similarities between habitats 3 and 5 in spite of the fact that habitat 4 separates the two (Fig. 2). This closer floristic similarity of habitat 5 with 3 is shown, for example, in the mosses, among which, in addition to the usual pioneers, three shade-mosses became established (Bryum argentum var. lanatum, Dicranum speirophyllum, and Thuidium plicatum). These arrived in year 9, indicating a change in microclimate near the ground at that time. This change was caused by the expanding crown-cover of the surviving Metrosideros trees. Other plants indicating similarities in microenvironment with habitat 3 were the grass Paspalum dilatatum, the sedges Cyperus polystachyos and Gahnia gahniaeformis, and the forbs Fragaria vesca var. alba, Cirsium vulgare, Anemone japonica, and Eupatorium riparium, which were common to only these two habitats (3 and 5).
It is interesting that no native woody plant invaded habitat 5. The only woody invaders were the exotic shrubs Rubus rosaefolius and R. penetrans.
Thin Fallout Area, Upper Kau Desert (Habitat 6)
This habitat had the least volcanic disturbance among the six. The pumice blanket varied from less than 25 cm two about 2 cm depth (Fig. 2). The habitat had been sparingly stocked with low-growing woody plants before the eruption. The scattered Metrosideros trees were scarcely more than 2-3 m tall. Recovery of surviving plants was of importance here, but a number of new invaders were recorded also.
Appendix VI shows the list of new invaders recorded in habitat 6. Algae arrived in year 2 and mosses, ferns, lichens, and seed plants (a small annual sedge, Bulbostylis capillaris) were added in year 3. Thus, seed plants arrived together with most other cryptogamic life forms in this habitat as in the others with snags or surviving plants. The early arrival of algae follows the general pattern observed in all other habitats. Thus, there is no significant departure in arrival sequence of life forms from the previous habitats, except two important pioneers were lacking. The moss Rhacomitrium lanuginosum var. pruinosum and the fern Nephrolepis exaltata did not appear among the pioneers. This is probably related to the lower rainfall. Another floristic difference is the appearance, among the pioneers, of the annual sedge B. capillaris, which is a characteristic plant on ash in the seasonal climate, but not found in the rain-forest climate (Doty and Mueller-Dombois 1966). The grass Rhynchelytrum repens also indicates the change to a drier climate in this habitat. The Rubus species did not invade; instead only two exotic woody plants (Psidium cattleianum and Buddleja asiatica) made their entrance in years 7 and 9 after the disturbance. As in habitat 5, Metrosideros tree seedlings did not appear in the 9-year observation period.
| <<< Previous | <<< Contents >>> | Next >>> |
chap5a.htm
Last Updated: 1-Apr-2005