|
SAGUARO
Ecology of the Saguaro: II NPS Scientific Monograph No. 8 |

|
CHAPTER 4:
ESTABLISHMENT AND SURVIVAL (continued)
Biotic Factors
Rodents and birds
Rodents are a major cause of first-year saguaro mortality. In the absence of other sources of food and/or moisture, most if not all of the species of rodents that occur commonly in the principal saguaro habitats at Saguaro National Monument and elsewhere in the Sonoran Desert will feed upon saguaro seedlings (Tables 13 and 14). In addition to direct destruction by consumption, a substantial number of young saguaros are uprooted or buried by intensive digging, especially by the Harris and round-tailed ground squirrels.
| Species | Seedlings consumed |
Digging activity (Soil disturbance) | |
| Hr | No. | ||
| Spermophilus tereticaudus | 17 | 56 | overturned |
| Citellus harrisi | 6 | 54 | overturned |
| Neotoma albigula | 5 | 65 | slight |
| Dipodomys merriami | 24 | 94 | overturned |
| Perognathus baileyi | 9 | 33 | overturned |
| Perognathus penicillatus | 24 | 24 | overturned |
| Peromyscus eremicus | 24 | 6.0 | none |
| Speciesa | Subspecies in area of | ||
| Southwestern Sonora (Thornscrub) |
Centwestern Arizona— Centwestern Sonora (Sonoran desertscrub) |
Northwestern Arizona (Mohave desertscrub) | |
Plant eaters | |||
| Desert Cottontail Sylvilagus auduboni |
a. goldmani | a. arizonae | a. arizonae |
| Antelope Jackrabbit Lepus alleni |
a. palitans | a. alleni | a. alleni |
| Blacktail Jackrabbit Lepus californicus |
c. eremicus | c. eremicus | |
| White-throat Woodrat Neotoma albigulab |
a. melanura | a. albigula | a. albigula |
| Sonoran Woodrat Neotoma phenax |
phenax | ||
| Desert Woodrat Neotoma lepidab |
l. devia | l. devia | |
| Cactus Mouse Peromyscus eremicus |
e. anthonyi | e. eremicus | e. eremicus |
| Riparian Mouse Peromyscus merriami |
m. goldmani | m. merriami | m. merriami |
| Deer Mouse Peromyscus maniculatus |
m. sonoriensis | m. sonoriensis | |
| Canyon Mouse Peromyscus crinitusb |
c. pallidissimus | c. pallidissimus | |
| Fulvous Harvest Mouse Reithrodontomys fulvescens |
f. tenuis | f. fulvescens | |
| Sonora Harvest Mouse Reithrodontomys burti |
burti | burti | |
| Western Harvest Mouse Reithrodontomys megalotis |
m. megalotis | m. megalotis | |
| Rock Squirrel Citellus variegatus |
v. grammurus | v. grammurus | v. grammurus |
| Roundtail Ground Squirrel Citellus tereticaudus |
t. neglectus | t. neglectus | t. neglectus |
| Sonora Antelope Squirrel Citellus harrisib |
h. harrisi | h. harrisi | |
| Desert Cliff Chipmunk Eutamias dorsalis |
dorsalis (Sonora) | ||
Seed gatherers | |||
| Merriam Kangaroo Rat Dipodomys merriamib |
m. merriami | m. merriami | m. merriami |
| Desert Kangaroo Rat Dipodomys desertib |
d. deserti | ||
| Bannertail Kangaroo Rat Dipodomys spectabilis |
s. perblandus | ||
| Ord Kangaroo Rat Dipodomys ordi |
o. ordi | o. chapmani | |
| Bajada Pocket Mouse Perognathus baileyib |
b. baileyi | b. baileyi | |
| Thornscrub Pocket Mouse Perognathus goldmani |
g. goldmani | ||
| Sinaloa Pocket Mouse Perognathus pernix |
p. pernix | ||
| Desert Pocket Mouse Perognathus penicillatusb |
p. pricei | p. penicillatus | |
| Rock Pocket Moose Perognathus intermediusb |
i. lithophilus | i. lithophilus | |
| Arizona Pocket Mouse Perognathus amplusb |
a. rotundus | a. pergracilis | |
| Little Pocket Mouse Perognathus longimembrisb |
I. bombycinus | ||
aScientific names, vernacular, and ranges for mammals in saguaro habitats follow Hall and Kelson (1959) the most recent reference, and communication from Drs. E. Lendell Cockrum (UA) and James L. Patton (UCB). bOther subspecies in area(s) in addition to tabulated. And other subspecies. | |||
We have found, however, that only the white-throated woodrat can subsist entirely on a diet of live saguaro tissues (Fig. 27). This is because the species of Neotoma are able to metabolize and neutralize oxalates, compounds that are lethal to other rodents. The story is well told by Schmidt-Nielsen (1964).

|
| Fig. 27A. Juvenile saguaro at Saguaro National Monument (east) severely damaged by white-throated woodrat (Neotoma albigula) and subsequent freezing. Woodrat tunnels often extend like a winding staircase completely through the stem. Such damage occurs on both juvenile and adult saguaros and increases vulnerability to freezing and to wind-caused stem breakage. Photographed 15 Sept. 1968. |
Seedlings of saguaros (and those of other cacti) that are not well hidden on the desert floor are vulnerable "canteens" that are approximately 90% water. In the desert, such cactus "feeding" rises sharply during the hot-dry months of May and June that precede the summer rains. Experimentally, however, we have found that under the hot-dry stress when ample free water and other fresh green plant material (grasses and forbs, both with and without seeds, and leaves) are available at the same time of year (and any other time), the same ground squirrels (Citellus) and desert mice (Peromyscus, Onychomys) refuse cactus of any species, age, or succulence. Because of oxalate poisoning, only one of these mammals (Neotoma) is able to incorporate the saguaro (and cacti in general) into its diet as a substantial food item. Desert rodents in general, and certain larger mammals as well (Fig. 27B), irregularly ingest limited amounts of cactus tissue primarily or wholly for its water content rather than for its caloric value. Packrats can and do have it both ways, saguaros included.

|
| Fig. 27B. Juvenile saguaro on the Cabeza Prieta Game Range (west of Organ Pipe Cactus National Monument). In the more arid portions of the saguaro's range, and elsewhere during periods of severe drought, jackrabbits utilize the succulent stem tissues as a source of moisture. Similar consumption by desert bighorn sheep has been observed (Simmons 1969). Although such damage does not in itself usually kill the saguaro, such "girdling" increases the plant's vulnerability to destruction by wind and freezing. Photographed 3 Feb. 1971. |
Destruction of recently emerged seedlings by cactus wrens has been observed. However, uprooted plants are not eaten. There is no doubt that the similar intensive foraging and digging activities of the curve-billed thrasher, gilded flicker, and gila woodpecker also result in destruction of young seedlings.
In nonrocky habitats, seedlings are rarely found near the base of adult saguaros, and almost never become established there. Such stations are subject to intense disturbance by foraging animals. Generally, the probability of seedling survival increases with distance from reproductive adult saguaros.
Insects
Young saguaros are subject to predation by a diversity of plant-eating insect species, none of which are known to feed exclusively on saguaros. The list of predators shown in Table 15 is by no means complete. Our own field observations and reports by others (Turner et al. 1966; Mann 1969) indicate that a complete listing of insect predators on young saguaros would include additional families, and many times the number of species identified. Although some insect-caused mortality of young saguaros takes place during every month of the year, the heaviest predation occurs during the humid summer months immediately following germination, and again with the arrival of warm weather in April and May.
| Species | Month collected | Stage |
| Lepidoptera | ||
| Cactobrosis fernaldialis (Hulst) | June, July | larva |
| Feltia subterranea (Fabricus) | June | larva |
| Heliothis zea (Boddie) | April, May | larva |
| Orthodes alfkeni (Grote) | July, Dec., Feb. | larva |
| Peridroma margaritosa (Hawthorn) | June, July | larva |
| Spodoptera exigua (Hubner) | April | larva |
| Orthoptera | ||
| Melanoplus sp. | July | nymph |
| Heleastus sp. | July | nymph |
| Gryllidae 2 spp. | July | adult |
| Coleoptera | ||
| Aneflus protensu (Le Conte) | July | adult |
During the first weeks following germination, the succulent seedlings are particularly vulnerable to destruction by the newly hatched larvae of lepidopterous insects. Immediately after hatching, the larva enters at the base or at the apex of the seedling to feed on the succulent inner tissues. Leaving the epidermis largely intact, the larva then moves on to an adjacent plant. The remaining withered epidermis presents the appearance of death resulting from lack of moisture. Some of the previously reported summer drought-kill based on observations of desiccated seedlings (Steenbergh and Lowe 1969) may be more correctly attributable to such destruction by insects. Subsequent observations indicate that insects are second only to rodents as a cause of seedling death during the first weeks following germination.
By the end of the first summer growth period, the relatively tough epidermal tissues protect the seedling from entry by small insect larvae and the well-developed spines protect the tender apex. However, the seedling remains vulnerable to destruction by larger insect predators: grasshoppers, crickets, beetles, and the larger larvae of certain moths.
Cutworms (noctuid moth larvae) appear to be the most common insect-consumers of saguaros that have survived beyond the first summer (Steenbergh and Lowe 1969; Table 15). Leaving only the roots and spines of demolished plants, cutworms frequently make a meal of several adjacent young plants (incidentally relieving competition for the survivors). Consumption of an entire 4-year-old plant (1.5 cm; 0.6 inch ht) within a 24-hr period was observed. The ability of the young saguaro to survive insect-caused damage increases with age and size—the saguaro outgrows the individual consumptive capacity of most predatory insects within the first 5 years of life.
Gerstaeckeria turbida (Lec.), a weevil reported by Turner et al. (1966) to be the principal insect responsible for the deaths of transplanted (cultured) young saguaros at Saguaro National Monument, was not observed during our investigations on causes of mortality in seedlings naturally germinated on-site in saguaro habitats.
A few species of large insects do occasionally invade larger juvenile saguaros. At Saguaro National Monument (east) the destruction of transplanted saguaros up to 30 cm (1 ft) tall by the larva of a large weevil [Cactophagus validus (LeConte)] has been observed. Our observations on naturally established saguaros, however, indicate that insect-caused deaths of such larger juvenile saguaros rarely occur in nature. Further, our observations strongly suggest that such destructive invasion of large juvenile saguaros by insects is limited to weak, moribund, or dead individuals damaged by freezing or other factors.
Experimental exclosures
Seed broadcasting and experimental exclosures were used on-site in both the east and west units of Saguaro National Monument (Tables 19-22; Figs. 19B, 29, 30). Details on the structure and placement of the wire exclosures are given in Steenbergh and Lowe (1976).
| Obs. date |
Elapsed days |
Survival | |||||||||||||
| East Monument |
West Monument | ||||||||||||||
| SEF N = 73 |
SERS N = 23 |
SERN N = 17 |
SEHS N = 10 |
SEHN N = 49 |
SWRS N = 26 |
SWRN N = 33 | |||||||||
| Live | % | Live | % | Live | % | Live | % | Live | % | Live | % | Live | % | ||
| 1967 | |||||||||||||||
| Aug. 1 | 31 | 9 | 39.1 | 10 | 58.8 | 7 | 70.0 | 8 | 16.3 | ||||||
| Aug. 2 | 32 | 19 | 73.1 | 29 | 87.9 | ||||||||||
| Aug. 3 | 33 | 10 | 13.7 | ||||||||||||
| Aug. 8 | 38 | 10 | 13.7 | 8 | 34.8 | 10 | 58.8 | 7 | 70.0 | 8 | 16.3 | ||||
| Aug. 9 | 39 | 17 | 65.4 | 23 | 69.7 | ||||||||||
| Aug. 15 | 45 | 10 | 13.7 | 8 | 34.8 | 10 | 58.8 | 3 | 30.0 | 8 | 16.3 | ||||
| Aug. 18 | 48 | 10 | 34.5 | 21 | 63.6 | ||||||||||
| Aug. 21 | 51 | 8 | 30.8 | 20 | 60.6 | ||||||||||
| Aug. 22 | 52 | 9 | 12.2 | 7 | 30.4 | 8 | 47.1 | 3 | 30.0 | 8 | 16.3 | ||||
| Sept. 7 | 68 | 5 | 19.2 | 13 | 39.4 | ||||||||||
| Sept. 8 | 69 | 4 | 5.5 | 6 | 26.1 | 6 | 35.3 | 3 | 30.0 | 4 | 8.2 | ||||
| Sept. 19 | 80 | 1 | 3.8 | 7 | 21.2 | ||||||||||
| Sept. 20 | 81 | 2 | 2.7 | 6 | 26.1 | 6 | 35.3 | 2 | 20.0 | 2 | 4.1 | ||||
| Oct. 3 | 94 | 1 | 3.8 | 1 | 3.0 | ||||||||||
| Oct. 4 | 95 | 2 | 2.7 | 5 | 21.7 | 1 | 5.9 | 1 | 10.0 | 0 | 0.0 | ||||
| Oct. 18 | 109 | 2 | 2.7 | 5 | 21.7 | 1 | 5.9 | 1 | 10.0 | 0 | 0.0 | 1 | 3.0 | ||
| Nov. 1 | 123 | 2 | 2.7 | 5 | 21.7 | 0 | 0.0 | 1 | 10.0 | 0 | 0.0 | ||||
| Nov. 15 | 137 | 1 | 1.4 | 5 | 21.7 | 1 | 10.0 | ||||||||
| Nov. 28 | 150 | 5 | 21.7 | 1 | 10.0 | ||||||||||
| Nov. 29 | 151 | 1 | 1.4 | 1 | 10.0 | ||||||||||
| Dec. 13 | 165 | 1 | 1.4 | 5 | 21.7 | 1 | 10.0 | ||||||||
| Dec. 19 | 171 | 1 | 1.4 | 5 | 21.7 | 1 | 10.0 | ||||||||
| 1968 | |||||||||||||||
| Jan. 4 | 187 | 1 | 1.4 | 3 | 13.0 | 1 | 10.0 | ||||||||
| Jan. 10 | 193 | 1 | 1.4 | 3 | 13.0 | 0 | 0.0 | ||||||||
| Jan. 24 | 207 | 1 | 1.4 | 3 | 13.0 | ||||||||||
| Feb. 7 | 221 | 3 | 13.0 | ||||||||||||
| Feb. 8 | 222 | 1 | 1.4 | ||||||||||||
| Feb. 21 | 235 | 1 | 1.4 | 3 | 13.0 | ||||||||||
| Mar. 6 | 249 | 0 | 0.0 | 3 | 13.0 | ||||||||||
| June 25 | 360 | 3 | 13.0 | ||||||||||||
| Habitat | N | Equation | r |
| SEF | 18 | Probit Y = -0.006 X + 3.897 | -0.882 |
| SEHS | 15 | Probit Y = -0.010 X + 5.225 | -0.828 |
| SEHN | 6 | Probit Y = -0.016 X + 4.645 | -0.922 |
| SERS | 2.0 | Probit Y = -0.003 X + 4.617 | -0.89.0 |
| SERN | 8 | Probit Y = -0.025 X + 6.230 | -0.943 |
| SWRS | 7 | Probit Y = -0.040 X + 6.762 | -0.968 |
| SWRN | 8 | Probit Y = -0.040 X + 7.299 | -0.982 |
| Habitat | N | Survivorship |
Establishment |
| To 0.001 (days) |
1st year (no.) | ||
| SEF | 73 | 334.6 | 0.4 |
| SEHS | 10 | 3269.0 | 0.1 |
| SEHN | 49 | 175.2 | <.0.1 |
| SERS | 23 | 1207.6 | 68.2 |
| SERN | 17 | 171.1 | <.0.1 |
| SWRS | 26 | 120.2 | <.0.1 |
| SWRN | 33 | 134.3 | <.0.1 |
|
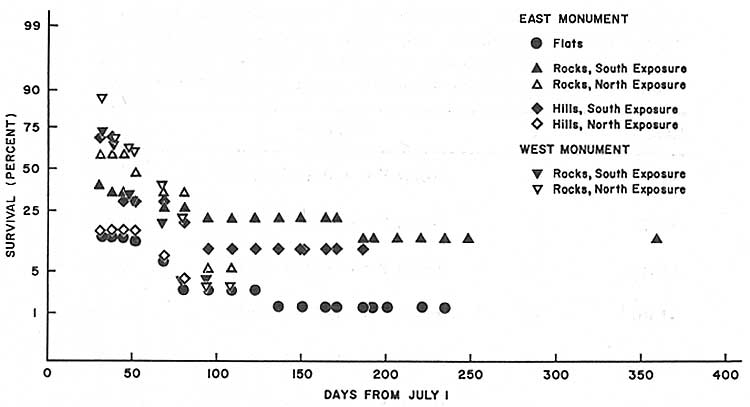
| Fig. 28. Post-germination first-year survivorship of saguaro seedlings (N = 231) naturally germinated July 1967 in representative habitats at Saguaro National Monument; percent survival on time in days from 1 July. Data in Table 16, regression equations in Table 17. (click on image for an enlargement in a new window) |
Comparison of 1968 seedling survival in open (unprotected) and exclosure (protected) plots clearly shows that vertebrate predators are a primary cause of saguaro seedling mortality during the first 5 pre-winter months of the first year of life (Tables 19-22; Figs. 29 and 30).
| Survival | |||||||||||||
| Obs. date |
Elapsed days |
East Monument |
West Monument | ||||||||||
| SEF N = 283 |
SERS N = 232 |
SERN N = 237 |
SWF N = 30 |
SWRS N = 151 |
SWRN N = 85 | ||||||||
| Live | % | Live | % | Live | % | Live | % | Live | % | Live | % | ||
| July 31 | 30 | 283 | 100.0 | 232 | 100.0 | 85 | 100.0 | ||||||
| Aug. 1 | 31 | 237 | 100.0 | 30 | 100.0 | 151 | 100.0 | ||||||
| Aug. 7 | 37 | 240 | 84.8 | 85 | 100.0 | ||||||||
| Aug. 8 | 38 | 221 | 95.3 | 227 | 95.8 | 30 | 100.0 | 145 | 96.0 | ||||
| Aug. 14 | 44 | 225 | 79.5 | 30 | 100.0 | 142 | 94.0 | 85 | 100.0 | ||||
| Aug. 15 | 45 | 206 | 88.8 | 185 | 78.1 | ||||||||
| Aug. 21 | 51 | 225 | 79.5 | 30 | 100.0 | 138 | 91.4 | 85 | 100.0 | ||||
| Aug. 22 | 52 | 155 | 68.8 | 161 | 67.9 | ||||||||
| Sept. 4 | 65 | 220 | 77.7 | 130 | 56.0 | 20 | 8.4 | ||||||
| Sept. 5 | 66 | 30 | 100.0 | 129 | 85.4 | 85 | 100.0 | ||||||
| Sept. 18 | 79 | 164 | 58.0 | 84 | 36.2 | 7 | 3.0 | ||||||
| Sept. 19 | 80 | 28 | 93.3 | 123 | 81.5 | 84 | 98.8 | ||||||
| Dec. 9 | 161 | 150 | 53.0 | 84 | 36.2 | 7 | 3.0 | ||||||
| Dec. 10 | 162 | 26 | 86.7 | 97 | 64.2 | 84 | 98.8 | ||||||
| 1969 | |||||||||||||
| Jan. 6 | 189 | 150 | 53.0 | ||||||||||
| Jan. 8 | 191 | 80 | 34.5 | 7 | 3.0 | 26 | 86.7 | 97 | 64.2 | 81 | 95.3 | ||
| Feb. 3 | 217 | 142 | 50.2 | 80 | 34.5 | 7 | 3.0 | 22 | 73.3 | 90 | 59.6 | 79 | 92.9 |
| Mar. 3 | 245 | 138 | 48.8 | 80 | 34.5 | 7 | 3.0 | 22 | 73.3 | 90 | 59.6 | 76 | 89.4 |
| April 1 | 274 | 138 | 48.8 | 80 | 34.5 | 7 | 3.0 | 22 | 73.3 | 90 | 59.6 | 75 | 88.2 |
| May 6 | 309 | 138 | 48.8 | 75 | 32.3 | 6 | 2.5 | ||||||
| May 7 | 310 | 22 | 73.3 | 89 | 58.9 | 75 | 88.2 | ||||||
| June 2 | 336 | 137 | 48.4 | 74 | 31.9 | 6 | 2.5 | 21 | 70.0 | 89 | 58.9 | 72 | 84.7 |
| July 1 | 365 | 137 | 48.4 | ||||||||||
| July 2 | 366 | 20 | 66.7 | 89 | 58.9 | 62 | 72.9 | ||||||
| July 3 | 367 | 74 | 31.9 | 6 | 2.5 | ||||||||
| July 31 | 395 | 137 | 48.4 | 74 | 31.9 | 5 | 2.1 | ||||||
| Aug. 1 | 396 | 18 | 60.0 | 79 | 52.3 | 57 | 67.1 | ||||||
| Survival | ||||||||||||
| Obs. date |
Elapsed days |
East Monument |
West Monument | |||||||||
| SEF N = 114 |
SERS N = 27 |
SERN N = 37 |
SWF N = 13 |
SWRN N = 59 | ||||||||
| Live | % | Live | % | Live | % | Live | % | Live | % | |||
| July 31 | 30 | 114 | 100.0 | 59 | 100.0 | |||||||
| Aug. 1 | 31 | 27 | 100.0 | 37 | 100.0 | 13 | 100.0 | |||||
| Aug. 7 | 37 | 94 | 82.5 | 58 | 98.3 | |||||||
| Aug. 8 | 38 | 22 | 81.5 | 30 | 81.1 | 11 | 84.6 | |||||
| Aug. 14 | 44 | 81 | 71.0 | 10 | 76.9 | 58 | 98.3 | |||||
| Aug. 15 | 45 | 9 | 33.3 | 23 | 62.2 | |||||||
| Aug. 21 | 51 | 25 | 21.9 | 10 | 76.9 | 43 | 72.9 | |||||
| Aug. 22 | 52 | 6 | 22.2 | 5 | 13.5 | |||||||
| Sept. 4 | 65 | 4 | 3.5 | 4 | 14.8 | 1 | 2.7 | |||||
| Sept. 5 | 66 | 5 | 38.5 | 26 | 44.1 | |||||||
| Sept. 18 | 79 | 0 | 0.0 | 1 | 3.7 | 0 | 0.0 | |||||
| Sept. 19 | 80 | 0 | 0.0 | 1 | 1.7 | |||||||
| Dec. 10 | 162 | 0 | 0.0 | 0 | 0.0 | |||||||
| Habitat | Survivorship and Establishment | |||||
| Open |
Exclosure | |||||
| N | To 0.001 (days) |
1st year (no.) |
N | To 0.001 (days) |
1st year (no.) | |
| SEF | 114 | 76.5 | <0.1 | 283 | 1442.0 | 498.6 |
| SERS | 27 | 100.2 | <0.1 | 232 | 968.8 | 223.4 |
| SERN | 37 | 74.2 | <0.1 | 237 | 462.9 | 60.6 |
| SE-(all) | 178 | 85.9 | <0.1 | 752 | 806.9 | 122.1 |
| SWF | 13 | 129.0 | <0.1 | 30 | 1316.4 | 629.0 |
| SWRN | 59 | 91.8 | <0.1 | 85 | 1001.7 | 737.8 |
| SWRS | — | 82.1a | <0.1a | 151 | 1128.4 | 463.1 |
aEstimated values based on analysis of 1967 data. | ||||||
| Habitat | N | Equation | r |
| UNPROTECTED PLOTS | |||
| SEF | 4 | Probit Y = -0.103 X + 9.805 | -0.979 |
| SERS | 5 | Probit Y = -0.056 X+ 7.471 | -0.920 |
| SERN | 4 | Probit Y = -0.109 X + 9.971 | -0.973 |
| SE (all) | 13 | Probit Y = -0.077 X + 8.558 | -0.837 |
| SWF | 4 | Probit Y = -0.046 X + 7.836 | -0.959 |
| SWRN | 5 | Probit Y = -0.100 X + 11.059 | -0.977 |
| PROTECTED PLOTS | |||
| SEF | 14 | Probit Y = -0.003 X + 5.788 | -0.846 |
| SERS | 14 | Probit Y = -0.004 X + 5.736 | -0.724 |
| SERN | 14 | Probit Y = -0.007 X + 5.108 | -0.701 |
| SE (all) | 42 | Probit Y = -0.004 X + 5.541 | -0.442 |
| SWF | 10 | Probit Y = -0.004 X + 6.675 | -0.941 |
| SWRS | 14 | Probit Y = -0.004 X + 6.423 | -0.884 |
| SWRN | 10 | Probit Y = -0.006 X + 7.870 | -0.962 |
| SW (all) | 34 | Probit Y = -0.005 X + 6.924 | -0.668 |
|
|
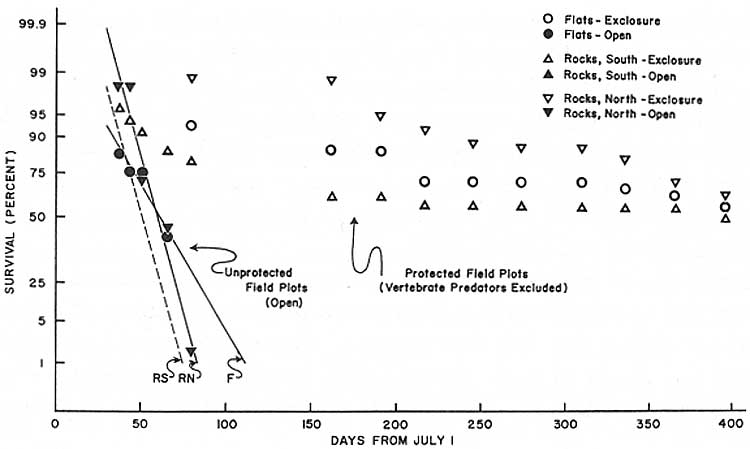
Fig. 29. First-year post-germination
survivorship in open and protected field plots of saguaro seedlings (N =
338) germinated July 1968 at Saguaro National Monument (west). As in the east monument, high rates of survival within exclosures indicate that mammals and birds are a primary cause of first-year seedling mortality in all habitats. Relative survivorship within exclosures on north-facing and south-facing slopes is reversed from that observed in the east monument (see Tables 21 and 22; Fig. 30). In this arid environment the higher rate of survival on the north-facing slope is attributed to the more favorable moisture relationships that prevail in that habitat during critical drought periods. Regression equations in Table 22, data in Tables 19 and 20. (click on image for an enlargement in a new window) |
The life expectancy of seedlings protected by exclosures that effectively excluded all vertebrate animals (but not insects) was approximately 10 times that of seedlings subject to natural predation (Table 21).
Within exclosures, consumption by insects was the principal cause of seedling deaths. However, despite relatively mild winter minimum temperatures, there were four freeze-caused seedling deaths within the two north-slope plots.
The relatively high rate of survival within the east monument "flats" exclosure is especially noteworthy. In that habitat, the pre-winter July to November climatic environment is highly favorable for seedling survival. There, also, it appears that the foraging activities of birds and rodents are most detrimental to seedling survival.
Although seedling survival in rocky habitats was improved vastly by the use of exclosures, the relative suitability of north-facing and south-facing slopes for seedling survival was not altered by the exclusion of vertebrate animals. The differences in the relative suitability of these habitats for pre-winter (July to November) seedling survival must be attributed to other factors, namely, consumption by insects and, to a lesser extent at Saguaro National Monument (west), to differences in moisture availability.
| <<< Previous | <<< Contents >>> | Next >>> |
chap4a.htm
Last Updated: 21-Oct-2005