|
SAGUARO
Ecology of the Saguaro: II NPS Scientific Monograph No. 8 |

|
CHAPTER 4:
ESTABLISHMENT AND SURVIVAL (continued)
Abiotic Factors
Drought
Drought-caused mortality of saguaro seedlings is related importantly to the date of germination and subsequent conditions for growth. Seedlings which sprout during July and early August usually attain sufficient size and volume during the summer growth period to survive through the autumn and fore-summer drought periods. During occasional years with heavy September and/or October rains, survival is further aided by additional growth.
Few seedlings that result from late summer germination (late August and September), however, survive to the start of the second year of life. Such seedlings do not develop sufficient water-storage capacity in the brief remaining growth period to survive prolonged periods of drought. Poorly developed seedlings that survive the arid after-summer are usually eliminated by the more severe drought conditions of the following arid fore-summer (Steenbergh and Lowe 1969). Our observations on winter mortality of seedling and young juvenile saguaros, however, support the conclusion that low levels of hydration increase resistance to freezing (Soule and Lowe 1970).
Desiccation and moisture stress resulting from freeze-caused injury to seedlings and young juvenile saguaros are important causes of apparent drought-kill during the arid fore-summer (Steenbergh and Lowe 1976). Although the death of such plants finally results from critical moisture stress, their death must be attributed to freezing rather than to drought.
Freezing
The results of the 1967 and 1968 seed-broadcasting experiments offer a reasonable measure of seedling establishment in the principal desert habitats at Saguaro National Monument during years with relatively mild winters (Tables 16-22; Figs. 28-30). During the two winters of those experiments, the lowest recorded minimum temperatures at the University of Arizona weather station were 22°F (-5.6°C) and 30deg;F (-11°C) 1deg;F (0.6°C) and 9°F (5.0°C) respectively, above the 77-year mean (21.1°F; -6.1°C) for that station.
|
|
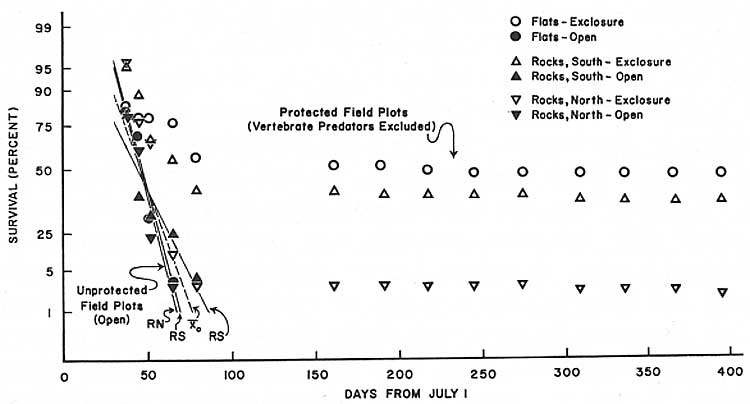
Fig. 30. First-year post-germination
survivorship in open and protected plots of saguaro seedlings (N = 930)
germinated July 1968 at Saguaro National Monument (east). The results clearly indicate that vertebrate predators (mammals and birds) are a primary cause of saguaro seedling mortality during the first year of life. In the flat (nonrocky) habitat, the rate of survival in the exclosure plot indicates that the pre-winter climatic environment in that habitat is highly favorable for seedling survival, and suggests that destructive foraging activities of birds and rodents are most intense in that habitat. Regression equations in Table 22, data in Tables 19 and 20. (click on image for an enlargement in a new window) |
The ultimate suitability of these habitats for the establishment and survival of young saguaros is determined, however, not by any climatic average but, rather, by the extreme minimum temperatures of the most severe winter that they must endure. Catastrophic freezes, a common occurrence in this region, critically limit survival not only of seedlings but of survivors from previous years as well (Steenbergh and Lowe 1976). At those times, the highest probability for continued survival of the young plant becomes abruptly and decisively associated with specific topographic situations (sensu microhabitats) that effectively moderate the intensity and duration of critical winter minimum temperatures, i.e., south-facing slopes, rocky footslopes, and the upper portions of adjoining valleys (Fig. 31). These are winter-warm microenvironments. Moreover, saguaros at the highest elevation (approximately 1585 m; 5200 ft) are growing only against south-facing (nocturnal re-radiating) rock outcrops (Fig. 32A). Conversely, in the colder microenvironments of north-facing slopes and areas of cold-air drainage and accumulation, there is a high rate of freeze-caused mortality; there is little or no survival of young saguaros in such habitats (Fig. 32B).

|
| Fig 31A. Sub-marginal habitat for saguaros—a north-facing slope overlooking the Cactus Forest at Saguaro National Monument (east). Saguaro seeds germinate, but young plants rarely become established in such habitats—most seedlings that survive through the warm pre-winter months die during the cold winter months of the first year of life. Thus few saguaros occur naturally on such sites. Cold air drainage from such north-facing slopes contributes to freezing conditions and resulting saguaro deaths in the Cactus Forest below. Photographed 21 Feb. 1969. |

|
| Fig. 31B. The warmer winter environment of this south-facing slope of the ridge, shown in Fig. 31A, offers more favorable conditions for saguaro establishment and survival than either the north-facing slope or the Cactus Forest habitat. However, this particular one is a marginal habitat for saguaros as evidenced by the freeze-caused constriction on the 2-m (6.6-ft) juvenile saguaro (left foreground) and the drooping top on the young adult plant (center right). The condition of the latter plant is the result of freezing at the site of a woodpecker nest-hole—a common cause of saguaro decapitation. Photographed 21 Feb. 1969. |

|
| Fig. 32A. Here at the upper limits of their elevational distribution (approximately 1524 m; 5000 ft) in Saguaro National Monument and in the nearby Santa Catalina Mountains up to 1585 m (5200 ft), saguaros grow against winter-warm south-facing (nocturnal re-radiating) rock outcrops. Saguaro National Monument photo files. |

|
| Fig. 32B. Freeze-killed 1.2-m (3.9-ft) juvenile saguaro on northeast-facing slope at Saguaro National Monument (east). The plant outgrew the adjacent rocks that protected it from freezing during the earlier years of its life. Photographed 21 Feb. 1969. |
On the upper bajadas (nonrocky, "flat" habitats), winter thermal microenvironments and freeze-caused mortality are intermediate between those of north-facing and south-facing slopes, and with increased distance from adjoining footslopes these habitats become progressively less favorable for the survival of young saguaros. Furthermore, as distance from the rocky footslopes increases, winter survival becomes increasingly dependent upon the presence of a multistoried (cold-moderating) canopy of associated shrubs and trees (Fig. 33).

|
| Fig. 33A. A forest of saguaros growing 53 km (33 miles) west of Sonoyta in northwestern Sonora, Mexico. Young saguaros here are associated with shrubs (primarily triangle bursage, Ambrosia deltoidea); note the absence of trees. In this relatively winter-warm locality, saguaros survive well without benefit of cold-moderating tree canopy. Here and in the few other flat habitats where it occurs in relative abundance, the saugaro is associated with sandy, well-drained soil. Photographed 21 Feb. 1972. |

|
|
Fig. 33B. A singularly outstanding
example of the association of young saguaros with "nurse-trees"—83
naturally established young saguaros are growing beneath this mesquite
tree (Prosopis velutina) in the Altar Valley, northern Sonora,
Mexico. The small shrubs are triangle bursage (Ambrosia
deltoidea). In nonrocky habitats in the colder portion of their distribution, asociation of saguaros with trees and large shrubs is intimately related to winter survival during periods of extreme cold. As the cold-limited boundaries of its range are approached, survival of the young saguaro is increasingly dependent upon the cold-moderating influence of a canopy of multistoried vegetation or massive outcroppings of rock. Photographed 18 Nov. 1969. |
Topography and slope exposure
Distinct differences in first-year saguaro survival occur in topographically different habitats of the two sections (east and west) of Saguaro National Monument. Field experiments comparing saguaro seedling survivorship in rocky habitats at the east monument clearly demonstrate that pre-winter survival on south-facing slopes is consistently higher than on adjacent north-facing slopes, and that the relationship continues throughout the first year of life (Tables 16-22; Figs. 28-30). Further, first-year survival in flat (nonrocky) habitats is lower than on south-facing slopes, but higher than on the north-facing slopes. Survival in the rolling-hills habitats is intermediate between that in flat and rocky habitats, and there, as on the steeper rocky slopes, seedling survival on the south-facing slopes is higher than on the north-facing slopes.
Due to lack of germination in some experimental plots, survivorship data from two west monument habitats are incomplete; in 1967, there was no germination at the "flats" site, and, in 1968, there was none in the open plot at the "rocks, south" site. However, synthesis of the available data for these years suggests that conditions for pre-winter seedling survival in the west monument habitats differ significantly from those of analogous habitats in the east monument. Data from the west monument (1967 experiments, and from 1968 exclosures) indicate that north-facing slopes offer a slightly more favorable environment for pre-winter survival of seedlings than do south-facing slopes. Compared to the east monument, the west monument offers a hotter and more arid environment. In that environment, relatively higher availability of moisture on north-facing slopes may well explain the higher rate of pre-winter survival in those habitats.
Soil moisture
The ratio of evaporation to soil moisture comprises a measurement of all the external factors which affect the waler relations of plants, except the influence of radiant energy on transpiration and the possible effects of soil temperature on this function. It is accordingly unnecessary to give further consideration to rainfall, which is not in itself a factor for vegetation, at least in such a region as Arizona.—Forrest Shreve, "The vegetation of a desert mountain range as conditioned by climatic factors." 1915:93.
Drought-caused mortality of saguaro seedlings, discussed previously in this chapter, refers to death from desiccation resulting from drying of the soil to beyond the wilting point. As also discussed, such mortality is related to details of topography, slope exposure, and other soil-related features of saguaro habitats.
The plant-available soil moisture is affected importantly by the physical texture of the soil (Table 23). Nowhere is the result of this fact more striking than in the Sonoran Desert home of the saguaro, for saguaros are not found in poorly aerated soils (Shreve 1920; Kramer 1961, 1962). Coarser-textured soils primarily of gravel and rock on bajadas, fans, and rock slopes support more highly diverse desert biotic communities in which the saguaro is a major dominant and where it often forms "saguaro forests" (Fig. 34A); such coarse soils are also relatively well drained. Finer-textured soils that are relatively high in clay, silt, and sand content support fewer species, and so result in desert communities of simpler structure (Fig. 34B); such relatively fine soils are also less well drained (Yang and Lowe 1956; Kramer 1962; Whittaker and Niering 1965).
| Soil characteristic | N | Larrea- Franseria | N | Cercidium- Cereus | t | P |
| Moisture equivalent (%) | 24 | 12.2 ± 0.61 | 24 | 6.6 ± 0.14 | 8.9 | <0.001 |
| Wilting coefficienet (%) | 24 | 6.6 ± 0.33 | 24 | 3.6 ± 0.08 | 8.8 | <0.001 |
| Moisture contenta (%) | ||||||
| Wet (approx. field capacity, summer rainfall season, July | 12 | 9.1 ± 0.90 | 12 | 5.5 ± 0.16 | 7.3 | <0.001 |
| Dry, post-summer drought season, December | 12 | 3.4 ± 0.41 | 12 | 1.4 ± 0.09 | 4.6 | <0.001 |

|
| Fig. 34A. The saguaro is codominant with foothill paloverde (Cercidium microphyllum) on this rocky southwest-facing slope in the Tucson Mountains, Saguaro National Monument. The cold-sensitive teddy bear cholla (Opuntia bigelovi) on the upper slope is an indicator of a winter-warm microclimate. Coarse-textured soils of gravel and rock bajadas, fans, and rock slopes support relatively diverse desert biotic communities in which the saguaro is a major dominant and where it often forms "saguaro forests." Photographed 28 April 1974. |

|
| Fig. 34B. Creosotebush community in the Avra Valley northwest of Tucson. The fine-textured soils of the valley floors that are relatively high in clay, silt, and sand content support relatively few species, and so result in communities of simpler structure than those on bajadas or rocky slopes as shown in Fig. 34A. The saguaro occurs rarely when at all on such poorly aerated soils. Photographed 28 April 1974. |
In experiments with young saguaros we have found that seedling growth stops and death eventually results from prolonged exposure to near-saturation soil moisture values in soils of all textures. In the greenhouse, rotting of young saguaros often results from daily watering during warm weather. In cultured young saguaros, Booth and Alcorn (1959) reported destruction of crowded plants by soft-rot fungi (Fusarium spp.).
The natural association of the saguaro with well-aerated soils and, conversely, its lack of tolerance of saturated soils under warm conditions strongly suggests that the southern limits of its distribution may be determined by interactions of soil characteristics and summer climate. The poorly aerated, predominately clayey soils that characterize the edge of the desert and coastal thornscrub southward from the vicinity north of Obregon to Navojoa may prevent the growth, establishment, and survival of young saguaros there and further southward in Mexico on the west Gulf Coast plain.
It is not surprising that the last saguaro populations southward are located on natural rock piles (Figs. 8, 12B, 35A). One is on Cerro Prieto east of Navojoa along the Navojoa-Alamos road and, further southward, the other is on Cerro Masiaca located northwest of Masiaca. Both populations are in extreme southern Sonora.

|
| Fig. 35A. Young saguaro established in a fractured basalt boulder at Cerro Masiaca in extreme southern Sonora. The two southernmost saguaro populations, at Cerro Masiaca and at Cerro Prieto (48 km = 30 miles northward), are located on boulder fields of rocky buttes well above and disjunctively isolated by the intervening level plains. Photographed 27 December 1974. |
As demanded by the hypothesis given, the habitats of both of these southernmost saguaro populations are on disjunctively isolated rocky buttes. These isolated populations are located well above the poorly aerated, fine-textured soils of silts and clays that characterize the intervening stretches of the flood-prone coastal plain. The intervening plain is densely clothed with subtropical thornscrub ("thorn forest") maintained by a relatively high summer-warm rainfall regime (Fig. 35B).

|
| Fig. 35B. Coastal thornscrub vegetation on the plains of northern Sinaloa: organpipe cactus (Cereus thurberi, left), hecho (Cereus pecten-aboriginum, center), morning-glory trees (Ipomoea arborescens, right), and dwarf jumping cholla (Opuntia fulgida mamillata, left foreground). The saguaro does not occur in these characteristic, poorly aerated, predominantly clayey soils of the thornscrub plains. The southern limits of saguaro distribution may be determined by its lack of tolerance of fine-textured and commonly saturated soils under warm conditions. Photographed 7 Jan. 1972. |
Factor
Interactions
In any year, the major portion of the current seedling crop is destroyed by a sequence of climatic and biotic factors. The operation of individual factors is associated specifically with one or more vegetative seasons as described by MacDougal (1908c, d) (Table 24): humid mid-summer (July to September), arid after-summer (October and November), winter months (December to March), and arid fore-summer (April to June). Most seedling deaths occur during the summer growth period. The rate of seedling mortality drops abruptly with the start of the arid after-summer in September (Tables 16-22; Figs. 28-30). Round-tailed ground squirrels estivate at that time, and insect numbers and activity decrease sharply.
| Period | Months | Term | Vegetation season (MacDougal 1908c) |
| I. The Basic Within-year Seasons and Terms | |||
| 1. July-Aug.-Sept. | 3 | Summer Rain | Humid Mid-summer |
| 2. Oct.-Nov. | 2 | Post-summer Drought | Dry After-summer |
| 3. Dec.-Jan.-Feb.-Mar. | 4 | Winter Freeze | Winter West Season |
| 4. April-May-June | 3 | Pre-summer Drought | Arid Fore-summer |
| II. Other Periods and Terms | |||
| 1. July-Nov. | 4 | Pre-winter | |
| 2. Dec.-Mar. (same as No. 3 above) | 4 | Winter | |
| 3. April-June (same as No. 4 above) | 3 | Pre-summer | |
There is large year-to-year variability in the rate of seedling establishment (Tables 25-27; Fig. 36). Such variability results primarily from differences in (1) summer rainfall and (2) winter minimum temperatures. During some years, exceptionally favorable conditions for germination produce an unusually large crop of seedlings; such conditions occurred at Saguaro National Monument (east) in 1964 (Steenbergh and Lowe 1969). Under those conditions many seeds germinate in exposed situations where there is little or no chance for seedling establishment. Under those circumstances a high rate of germination will be followed by high seedling mortality. Thus, a high rate of seed germination does not necessarily result in a correspondingly high rate of seedling establishment.
| Year | Plots N |
Seeds broadcast |
Total germination |
Survivorship observer | |||
| N | % | N | % | Base (seeds) | |||
| 1965 | 28 | 28,000 ± 50 | 96 | 0.34 | 96 | 100.0 | 28,000 |
| 1966 | 6 | 19,000 ± 50 | 74 | 0.39 | 73 | 98.65 | 18,743 |
| 1967 | 19 | 95,000 ± 100 | 459 | 0.48 | 328 | 71.46 | 67,887 |
| Total | 53 | 142,000 ± 100 | 629 | — | 497 | — | 114,630 |
aIn 1967 there could be no observations on survivorship in 7 of the 13 plots due to lack of germination. | |||||||
| Date | 1965 (N = 96) |
1966 (N = 73) |
1967 (N = 328) | ||||||
| Cum. dead | Surv. N | Surv./million seeds |
Cum. dead | Surv. N | Surv./million seeds |
Cum. dead | Surv. N | Surv./million seeds | |
| Aug. 1 | 153 | 175 | 2,578 | ||||||
| Aug. 6 | 3 | 70 | 3,735 | ||||||
| Aug. 8 | 178 | 150 | 2,210 | ||||||
| Aug. 11 | 23 | 73 | 1,607 | ||||||
| Aug. 13 | 12 | 61 | 3,255 | ||||||
| Aug. 15 | 203 | 125 | 1,841 | ||||||
| Aug. 18 | 59 | 37 | 1,321 | ||||||
| Aug. 20 | 21 | 52 | 2,774 | ||||||
| Aug. 21 | 213 | 115 | 1,694 | ||||||
| Aug. 26 | 79 | 17 | 607 | ||||||
| Aug. 27 | 31 | 42 | 2,241 | ||||||
| Sept. 1 | 89 | 7 | 250 | ||||||
| Sept. 3 | 35 | 38 | 2,027 | ||||||
| Sept. 7 | 247 | 81 | 1,119 | ||||||
| Sept. 9 | 91 | 5 | 179 | ||||||
| Sept. 10 | 36 | 37 | 1,974 | ||||||
| Sept. 13 | 91 | 5 | 179 | ||||||
| Sept. 17 | 37 | 36 | 1,921 | ||||||
| Sept. 19 | 275 | 53 | 781 | ||||||
| Sept. 22 | 92 | 4 | 143 | ||||||
| Sept. 25 | 44 | 29 | 1,547 | ||||||
| Oct. 2 | 48 | 25 | 1,334 | ||||||
| Oct. 3 | 303 | 25 | 368 | ||||||
| Oct. 6 | 92 | 4 | 143 | ||||||
| Oct. 18 | 306 | 22 | 324 | ||||||
| Oct. 23 | 96 | 0 | — | 50 | 23 | 1,227 | |||
| Oct. 30 | 53 | 20 | 1,067 | ||||||
| Nov. 1 | 53 | 20 | 1,067 | 314 | 14 | 206 | |||
| Nov. 6 | 53 | 20 | 1,067 | ||||||
| Nov. 13 | 54 | 19 | 1,014 | ||||||
| Nov. 15 | 317 | 11 | 162 | ||||||
| Nov. 29 | 319 | 9 | 133 | ||||||
| Dec. 11 | 55 | 18 | 960 | ||||||
| Dec. 13 | 320 | 8 | 118 | ||||||
| Dec. 19 | 320 | 8 | 118 | ||||||
| Jan. 4 | 322 | 6 | 88 | ||||||
| Jan. 8 | 60 | 13 | 694 | ||||||
| Jan. 10 | 323 | 5 | 74 | ||||||
| Feb. 5 | 62 | 11 | 587 | ||||||
| Feb. 8 | 324 | 4 | 59 | ||||||
| Feb. 21 | 324 | 4 | 59 | ||||||
| Mar. 5 | 66 | 7 | 373 | ||||||
| Mar. 6 | 325 | 3 | 44 | ||||||
| Apr. 4 | 67 | 6 | 320 | ||||||
| Apr. 30 | 68 | 5 | 267 | ||||||
| May 29 | 71 | 2 | 107 | ||||||
| June 26 | 96 | 0 | 72 | 1 | 53 | 325 | 3 | 44 | |
| Year | N | Equation | r |
| 1965 | 8 | log Y = -3.294 log X + 8.498 | -0.946 |
| 1966 | 20 | log Y = -1.464 log X + 6.003 | -0.933 |
| 1967 | 17 | log Y = -2.012 log X + 6.563 | -0.985 |
| Pooled | 45 | log Y = -6.078 log X + 6.244 | -0.939 |
|
|
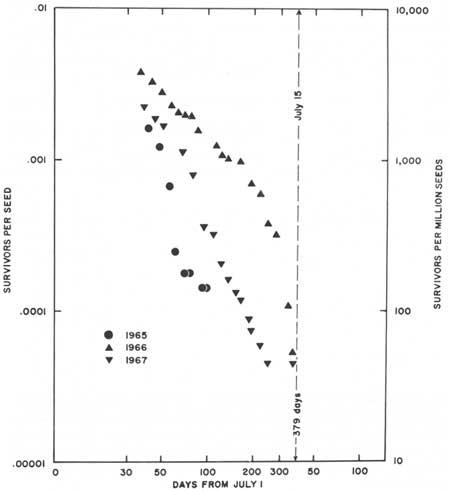
Fig. 36. Net first-year survivorship
(from seeds) for 497 saguaro seedlings at Saguaro National Monument.
Survivorship from 1 July is shown as seedling survivors per million
seeds broadcast in representative natural habitas (Table 25); least
squares regression of survivors on time in days. Based on these data obtained during 3 consecutive years from July 1965 to June 1968, the estimated mean establishment rate (i.e., survival to 15 July of the following summer, the start of the second year of life) for saguaros in the Tucson area is 51.0 plants per million seeds (0.005%). The estimated post-germination establishment rate is 1.26%. Regression equation in Table 27, data in Tables 25 and 26. |
The relative importance of each of the several factors responsible for the destruction of young saguaros cannot be measured simply in terms of the relative mortality attributable to each cause. Rather, the relative importance of each factor is time-related. The importance of a particular factor lies in both its temporal and spatial impact upon the existing population and the life expectancy of the survivors. Furthermore, evaluation of factor importance must be concerned with the question of whether or not the particular factor operating is density-dependent and age-related.
The life expectancy of a young saguaro increases rapidly with age—it outgrows its predators and becomes increasingly able to survive the periodic hazards of the abiotic environment. However, in the case of recurring catastrophic freezes which operate without regard to population density (density-independent), survival is primarily dependent upon (1) the number of winter-favorable sites available within the habitats, and (2) the number of such sites actually occupied by young plants. Regardless of their numbers, other exposed cohorts in the population will die. Thus, near the cold-limited boundaries of its range, the number and character of physical microhabitats, as determined by the plant community together with the topographic characteristics of the habitats, are the factors that ultimately limit the survival and density of young saguaros.
| <<< Previous | <<< Contents >>> | Next >>> |
chap4b.htm
Last Updated: 21-Oct-2005