|
NATIONAL PARK SERVICE
The Giant Sequoia of the Sierra Nevada |

|
CHAPTER 5:
LIFE HISTORY
Reproduction
To continue as a living species, a higher plant must succeed in community competition at the most precarious point of its life history, which for land plants, at least, is usually the period of reproduction. A plant's reproductive success requires the following exacting sequence during this period:
1. The production of sufficient viable seeds.
2. A dependable method of seed dispersal.
3. A suitable substrate for germination.
4. Climatic conditions conducive to germination of the seeds.
5. Adequate light and soil moisture for seedling survival.
6. Survival of enough seedlings to maintain the species.
The responses of plants to environmental factors vary greatly with the genetic make-up of the species; the more exacting these requirements and their timing, the more tenuous the plants' reproductive success.
The giant sequoia is a classic example of reproductive fragility in the plant world, eloquently expressed in its restricted and much interrupted natural range, the relative stability of grove boundaries, and the diverse age-class structure within individual groves. Many of the groves have produced little or no progeny within the relatively short period since the beginning of western civilization in California. The much more extensive range of sequoian ancestors has led several persons to believe that the species may be nearing extinction, much like the dinosaurs of the past. Here rests an interesting parallel: the sequoia and the dinosaur represent the largest developments of a land organism within its kingdom in the earth's history.
One of the most misunderstood facets in the sequoia life cycle is that of its reproductive requirements and the sequence of events which lead to the successful perpetuation of its kind. Until now the story, fragmented and erroneous, has been passed without question from one generation to the next by amateur and scientist alike. Only recently, following an extensive series of objective studies, have the ecological relationships of the sequoia life cycle begun to be well understood.
Cone and seed production
The reproductive sequence generally begins in mid- to late winter, when the tiny staminate (male) cones literally cover the outer branchlets of the crown. At the height of pollen dispersal, golden clouds of this pollen or male reproductive cells may be seen drifting about on the slightest breeze or staining the snow with their yellowish tint. This great abundance of pollen simply ensures reproduction, other conditions permitting. Such safety in numbers is typical of organisms faced with low reproductive success.
At the time of pollination, the female cone is only about the size of a grain of wheat and is hard, with a pearly gray tint (Fry and White 1930). In the first summer after fertilization, the developing cones begin to produce chlorophyll, which colors them bright green. At the end of the first growing season, the cones are usually more than three-quarters their full size and the cone scales are very soft and fleshy. From the flattened apices of the scales project slender hair-like bracts, which identify the cones as immature. The seeds, including the area occupied by the embryo, are now a light straw color, and generally cannot germinate—although, in an experiment at San Jose State University, a single first-year seed successfully germinated on wetted filter paper in a petri dish. Because first-year seeds have little opportunity to be disseminated onto the ground, their role in regeneration is very much limited. At the end of their second growing season, the cones attain maturity and generally produce viable seeds.
By late summer of the second growing season, the cones approximate their full size. The cone scales are somewhat woodier in texture the hair-like bracts will have now fallen away, and the cone is usually a dark forest green. Maturity of the seeds is clearly indicated by the dark brown, longitudinal stripe which runs through the flat oval wing and marks the place where the embryo lies.
With maturity, sequoia cones behave differently from those of most other conifers, neither turning brown nor in any way commencing to disseminate their seeds. Rather, the cones remain attached to the stems in a green, active photosynthetic state, and they increase slightly in size each year so that the cone scales, becoming bulbous, give the older cones a rather knobby appearance. There is good evidence that the vascular connections between the cones and the seeds remain intact as long as the cones remain green on the trees. Cones remain in this state rather commonly for 8-12 years and at least one green cone was determined to be 22 years of age by Buchholz (1938). After 4 or 5 years, cones may begin to support a growth of foliose lichens, which sometimes completely cover them (Fig. 22).
We determine the age of sequoia cones just as we determine the age of a tree's trunk, namely, by counting the annual growth rings in the peduncle or stem of the cone. A clear cross-sectioned cut with a razorblade or knife will suffice to expose the rings for counting. The first 2 years' rings are large enough to be seen with only slight magnification, but the later ones are often extremely narrow and difficult to distinguish (Fig. 31).
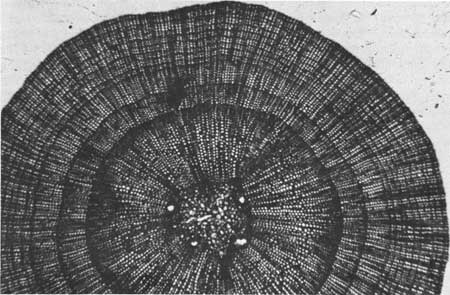
|
| Fig. 31. Cross section of sequoia cone peduncle showing annual rings. At maturity, sequoia cones do not open and shed their seeds. They are retained as living entities for as much as 21 years. Age of living cones may be determined by counting the annual rings in the peduncle. Photo by R. J. Hartesveldt. |
The phenomenon of green cone retention poses some interesting questions. Perhaps foremost is the situation in which mature, viable seeds are retained in a moist cone where summer temperatures are surely within the range required for germination. What, then, prohibits germination of the seeds in these circumstances? Experiments indicate that it may well be the function of the reddish, crystalline-like substance found between the cone scales. This substance, often referred to as red cone pigment, is in a liquid form at the time the cones are green. It is an amorphous, water-soluble compound, like the blackened exudate found on scarred parts of sequoia trunks, and makes up about half of the seeds' weight. The function of this pigment has been much debated over the years, in part because green cone retention was not understood. In the early part of the century, analyses indicated that the pigment was very high in tannin, and therefore many assumed that its role was to prevent insect and fungus attack. Fry and White (1930) felt that the pigment helped maintain the seeds' viability over the years. Beetham (1962), after soaking the seeds in a solution of cone pigment for 2 months, found no apparent advantage or disadvantage to seed viability or to the resulting seedling. She did find that planted seeds which had been treated with an 85% solution for 2 months were very slow to germinate, but that the total final germination was very near that of the controls.
Martin (1957-58) obtained a rather different result from similar experiments in northern Germany. Although his studies concur with Beetham's in showing that seed germination time increased with increased concentration of the cone pigment extract, he found that in concentrations of 30% and more the seeds failed to sprout at all. Thus, he reasoned that the pigment's role was to arrest germination of the seeds within the cones through the establishment of a reverse osmotic gradient. He further reasoned that when the cones opened, the liberation of the seeds occurred only when rainfall had dissolved and washed away the cone pigment.
Martin reports identification of the pigment chemical as tannin glucoside (C21H20O10). In fungus cultures treated with the extract, however, not even its highest concentrations inhibited their growth. Such results do not necessarily negate the possible role of the pigment as a fungal preventive. Recent studies also indicate that at least two species of insects feed regularly within sequoia cones, the pigment apparently not deterring them in the slightest. Clearly, further investigation is necessary to clarify the exact role of the pigmenting chemical.
Because the pigment is soluble in water, naturalist John Muir made ink of it and wrote numerous letters with the fluid. Letters written 60-70 years ago are still clearly legible today. The Forest Products Laboratory in Madison, Wis., analyzed this substance for its potential as commercial ink, but samples, submitted to the Carter Ink Co., clogged pens readily because of gummy accumulations that attended the evaporation of the water (Kressman 1911); thus nothing ever came of it.
The other major question in cone retention is over just what finally triggers the browning of the cones to permit seed dispersal. Ring counts on the penduncles of browned cones removed from living trees show that the drying process usually commences after the 5th or 6th year of the cone's life on the tree, although it may often start only several years later. This great variation among individual trees largely eliminates genetic time-switches and climatic variations as causative factors. Stecker (1969) discovered that cone drying resulted from the activities of a small cerambycid beetle, Phymatodes nitidus, which feeds upon the cone flesh. This subject will be considered in greater detail under "Seed Dispersal and Cone Fall."
Beginning age of cone production
Estimates of the age at which sequoias begin to produce cones also vary greatly in the literature, perhaps because authors have used different standards. While some have given the species' average age at which cones begin to appear, others have been specific for individual trees, and some give the age of abundant cone production. Still others cite the time at which cones bearing viable seeds are produced. For instance, Harwell (1947) sets an age of 200 years for the beginning of cone production. Clarke (1954) gives 175-200 years; the U.S. Department of Agriculture (1948), 125 years; Cook (1955), 70 years; Metcalf (1948), 50 years; Wulff et al. (1911), 24 years; and Anon. (1960), 11 months (Fig. 32). This represents a ratio of estimate of 218:1.

|
| Fig. 32. Sequoia cone on 11-month old potted specimen, Argonne National Laboratory, Illinois. Perhaps the youngest sequoia ever to produce a cone, this specimen was subjected to an abnormally long photoperiod of 16 hours of light per day. Although fairly common on natural specimens in the field, these cones seldom attain maturity and are assumed to produce few if any viable seeds. Photo courtesy of Argonne National Laboratory. |
This great reported variation is interesting because we have a long record of sequoias that have borne cones at an early age. The first such reports came out of England, where proud gardeners probably watched their precious newly imported specimens with intense interest. The Gardener's Chronicle (London) (Anon. 1859) reported a 4.5-ft specimen of Wellingtonia bearing cones, and Muggleton (1859) records a cone on a 36-inch specimen. Numerous other reports of cones borne at an early age came from throughout northern Europe in the following decade. None of these earliest cones bore viable seeds, however, which raised considerable curiosity until it was discovered that sequoias begin to produce ovulate cones several years before staminate cones, so that pollen was not at first available for fertilization in Europe. When staminate cones eventually did begin to develop, so did viable seed. Brown (1868) records fertile seed produced for the first time in England on a specimen that he felt could not have been more than 10 years old. Undoubtedly, some of the cones removed for seed viability tests were first-year cones which bore only immature seeds.
We have experimented with seeds taken from trees estimated to be within 10- to 14-year range that had been planted by the U.S. Forest Service on the McGee Burn in the Converse Basin of Sequoia National Forest. Cones removed from near the tops of several 12- to 15-ft specimens yielded a total of 220 seeds. These were placed in petri dishes for germination. A total of 64 seeds, or 29%, germinated, a viability percentage which compares favorably to that of seeds taken from trees several hundred years old. (Hartesveldt et al. 1967).
The specimen recorded by Anon. (1960) as producing a cone at 11 months was perhaps influenced by unnatural growth conditions at the Argonne National Laboratory in Illinois. It was subjected to an abnormally long daily photoperiod of 16 hours for its entire 11 months, plus a controlled temperature regime. The seedling was only 16 inches tall when its leader shoot began to swell and formed a cone which eventually arrested the growth of the terminal shoot. A lateral branch took over the leader shoot role, and the cone and its shoot became lateral. While this specimen was unusually young and small for cone production, such cones are not uncommon in nature on trees 6-10 ft tall and perhaps 8-10 years of age. The cone's tip usually bears the continuation of the leader shoot. Although tests have not been made, it has been generally assumed that these cones bear no viable seeds.
Sequoias of all sizes and presumably of all ages are prolific bearers of cones and viable seeds. We know of no age beyond which the tree's reproductive capacity stops or even diminishes. Even severely damaged trees continue to bear abundantly.
Cone abundance
Because the crown is so high and the foliage so dense, it has been difficult at best to obtain reliable estimates of the total number of cones on mature trees. Recently, with an elevator rigged in a 290-foot specimen now known as the Castro Tree in the Redwood Mountain Grove of Kings Canyon National Park, Steckler has made in-crown studies which may now permit more accurate estimates than those made from the ground by Shellhammer. These studies indicate that previous estimates of cone-load and number of seeds dispersed yearly have been greatly in error. Almost certainly, earlier authors have also misunderstood the life span of living green cones and the ratio of green cones with viable seeds to browned cones with none.
According to these in-crown studies, a large sequoia tree might be expected to contain at any given time about 11,000 cones, of which perhaps 7000 would be closed, fleshy, and photosynthetically active. The remaining 4000 cones would be opened, brown, and largely seedless, although Fry and White (1930) record viable seeds remaining in cones 16 years after the latter have turned brown. An average of 1500-2000 new cones are produced in the average year. Occasionally, optimal weather conditions greatly increase the production of new cones. For instance, in 1970, the Castro study tree produced 20,697 new cones. Each year somewhat fewer older cones, both green and brown, are felled by wind, rain, and snow than the previous year. Variations in total cone-load are very probably genetically controlled, and certainly controlled by site location; very large specimens growing on favorable sites may bear more than 40,000 cones at one time, while those on the poorer sites may have as few as 6000. The upper part of the crown of any mature tree invariably produces a greater abundance of cones than its lower portions (Fig. 33).

|
| Fig. 33. Diagrammatic view of cone-load distribution in m giant sequoia. Drawing by Win Stiles. |
A correlation apparently exists between the number of new cones produced each year and the quantity of available soil moisture during the winter and spring months. The cones produced in wet years are more numerous and yield seeds of greater viability than those produced in dry years.
Seed viability
While Fry and White (1930) record the average viability of sequoia seeds as only 15% of the total crop, our experience shows that 35% is a more representative figure. The actual percentage of viable seeds produced varies with such factors as the tree's topographic site, the cone's age, its specific location in the crown, and the seed's position within the cone. Large seeds usually are more viable than small ones.
Initial studies show that rocky slopes and ridges are definitely more advantageous for seed viability than flattish bottom lands with deeper soils. Our experimental plots in the Redwood Mountain Grove have trees on the rockier slopes yielding seeds with 54% average viability, whereas in the flats along Redwood Creek the figure is 32%.
In preliminary studies with cones of various ages, it was found that seeds which retain their vascular connections with the cone continue to grow. This is partially borne out by the viability statistics in Table 2 derived from both snap tests and actual germination tests in petri dishes. The snap test involves breaking the seed across the embryo and noting its color. If it is an off-white color and the embryo completely fills the embryo case, it is most likely viable. However, if it is snow-white and shriveled, or brownish, the seed is not viable (Clark 1907).
On the basis of a previous assumption that the seeds of the living cones grow and perhaps increase in size and viability, the decrease in viability following the fifth year appears inconsistent. However, the beetle-feeding activities mentioned earlier provide a plausible explanation.
TABLE 2. Germination for sequoia seeds of various ages.
| Age of seeds and cones | Viability by snap test | Viability by germination |
| 2 years | 26% | 20% |
| 3 years | 25% | 32% |
| 4 years | 27% | 33% |
| 5 years | 50% | 52% |
| 6 years | 32% | 41% |
| 7 years | 25% | 32% |
| 8 years | 34% | 27% |
Within individual cones, the seeds' germination was 26% in the basal portion (nearest the peduncle), 59% in the central portion, and 36% in the apical region. Perhaps this variation is due to effective vascular connection between the seeds and cone scales, which in turn may also control the size of the seeds produced. This, however, is yet to be verified.
Studies indicate that cones produced near the tops of the trees tend to be smaller than those lower in the crown, and that the larger cones produce a higher percentage of viable seeds. Metcalf (1948) records large cones with seeds showing a 75% viability, a figure which Stecker has verified in cones from the crown of the Castro study tree.
The older cones cluster along the branches' main axes, back from the growing tips; the newest cones are in groups of 2-19 at the very ends of the main branches and are often surrounded by dense new foliage; the mature cones of various age classes are distributed proportionately between these two extremes. Cones from the last 3- or 4-year age groups, that is, ages 1 through 3 or 4, are about equally represented on a branch. About 65% of the living cones are up to 5 years old, 25% are 6-10 years old, and up to about 10% are 11-20 years old.
Seed dispersal and cone fall
Over the years, writers have quoted others who said that a mature sequoia tree sheds close to a million seeds a year, a figure derived by unknown methods and which the in-crown studies show to be a gross overestimate. Ground-level calculations, however, were undoubtedly difficult because of the dense foliage obscuring many of the inner cones. Certain assumptions are necessary to obtain a more realistic figure of annual production.
We can assume that the average number of cones opened or otherwise lost per year probably equals the average number produced. Then, according to the given average of 1500-2000 new cones per year and an equal loss due to browning and falling, and if we assume 200 seeds per cone (see page 88), the mature sequoia would disperse between 300,000 and 400,000 seeds per year. This means a potential seed dispersal of from 200,000 per acre per year to perhaps as many as 2 million.
Dispersal takes two different forms: seeds may be distributed from tree-top level as the cones either open upon browning or are eaten by chickarees on the limbs of the crown; or, the cones may be cut or otherwise fall to the ground, where they dry out and spill their seeds upon a relatively small area of the soil or leaf litter surface. Each has advantages and disadvantages. A very important point here is that a high percentage of sequoia seed is dispersed by the activities of animals.
The chickaree (Fig. 34), inhabits many trees in the Sierran forest region. Outside sequoia groves, the chickaree feeds commonly on the seeds of the sugar pine, white fir, red fir, ponderosa pine, incense-cedar, etc. But in sequoia groves where sequoias displace some of the other tree species, this handsome little squirrel also feeds on the fleshy green scales of the younger sequoia cones, much as people do in eating the flesh from artichoke bracts. In the process, the seeds, too small to have much food value for the squirrel, are dislodged and, if the cone is eaten on a high limb, they are scattered over the ground. Dispersal from this height has the potential advantage that wind drift may extend the species' range.

|
| Fig. 34. The chickaree, or Douglas squirrel, is now believed to play a significant role in the reproduction of the giant sequoia. This small rodent is apparently more interested in the flesh of the cone scales as a source of food than the tiny seeds. In the process of feeding upon the cone scales, many seeds are spread upon the soil surface. Photo by Tom Harvey. |
In circumstances not fully understood, chickarees will cut innumerable cones from individual trees, dropping them immediately to the ground. They may cut cones from individual trees for 6 or 7 years in a row, thus changing the cone-load factor for a considerable period. Shellhammer (1966) observed a lone chickaree which cut down 539 green sequoia cones in 31 minutes. There are many records of large cone cuttings in a single day or during a season, one of the most amazing being that of Fry and White (1930). In 1905 they recorded a single chickaree cutting cones which, when gathered up, filled 38 barley sacks and yielded 26 lb of seeds. Calculating 91,000 seeds to the pound and 200 seeds per cone, we obtain a figure of nearly 12,000 cones.
Once the cones are cut, the chickaree caches them away as future food. Because the cones are edible only when green and tender, the storing must be such as to maintain this condition over the longest possible time. Many chickarees will bury the cones individually in the leaf litter and duff, or in caches of six or seven cones where space permits. In places, often in the bed of an intermittent stream where the soil is moist or bog-like, they will store hundreds or even thousands of cones in impressive piles. Sometimes the space between a fallen sequoia log and the ground is used, or the base of a hollow tree may be packed tightly with cones. The chickaree, returning in fall and winter to eat, usually spills the seeds onto the ground in the relatively small space where it feeds. This wastage certainly contributes to the sequoia's potential regeneration, although without the advantage of wind dispersal or of shallow burial in soft, friable soil following a fire. However, seeds properly placed during this feeding process are in good cold storage throughout the winter and so are ready for spring germination. The many cones not eaten by chickarees eventually dry out and spill their seed contents too.
In late summer and autumn, especially on warm days with a breeze, sequoia seeds flutter to the ground in an almost constant rain. Their small oval wings help insure that the slightest movement of air will carry them away from the parent tree, perhaps as much as 600 ft. The already mentioned feeding activities of the beetle, Phymatodes nitidus (Fig. 35), causing the cones to turn brown, probably bring about much of this seed fall.
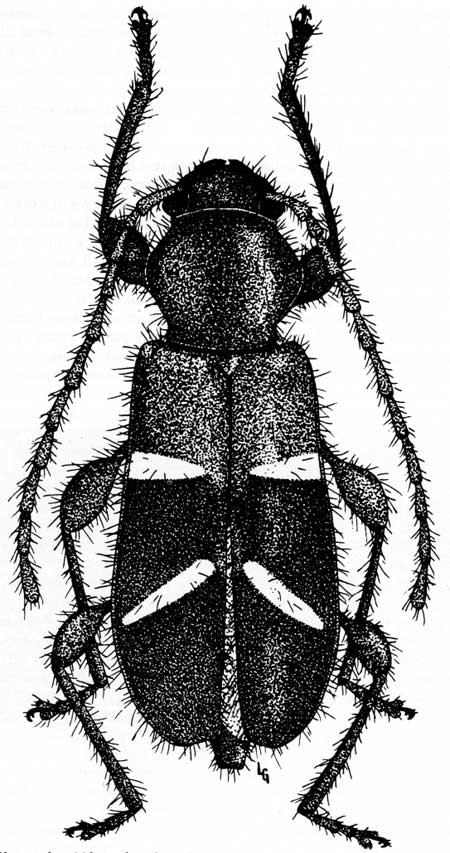
|
| Fig. 35. Phymatodes nitidus, a long-horned, wood-boring beetle, responsible for dissemination of some of the crop of sequoia seeds from tree crown levels. The larvae of these beetles feed on the interior flesh of green cones and eventually sever enough vascular connections to kill the cone. This results in the opening of the cone and eventual dispersal of the seeds. Drawing by Loren Green. |
Stecker (1969) discovered this beetle high in the crown of the Castro Tree during the summer of 1968. In virtually all the browned and dried cones, tiny insect emergence-holes were found. These insignificant indications of insect activity, having somehow eluded discovery by science until then, gave the first evidence that browning was not simply a natural aging of the giant sequoia's cone tissues. Dissection of thousands of these cones showed that the agent is a very small cerambycid or long-horned wood-boring beetle larva (Phymatodes nitidus LeC) and, furthermore, that it plays an important role in the giant sequoia's regeneration. The larvae's length is only 3-5 mm (1/8 to 1/5 inch).
Oviposition by the female beetle takes place at cone-scale junctures and occasionally along the cone stem or peduncle. Upon hatching, the larval borer chews its way into the cone's interior, obtaining nourishment from the carbohydrate-laden tissues of the cone scales. The mines are about one-third the diameter of a lead pencil and are packed with chewing and digestive waste, or frass, which in this case resembles a fine salt-and-pepper mixture. The vascular channelways or veins (two layers of them in each cone scale of giant sequoia) are often severed during this feeding, which diminishes water conduction to the ends of the cone scales. At first one scale will brown, and then others, as they dry out, following the direction taken by the feeding larvae.
As the mining turns the green, fleshy tissues into a sun- and air-dried cone, the flesh shrinks, creating gaps between the cone scales. And so, the cone's hold upon the seeds is relaxed and dispersal follows. Seeds thus released from the tree's top, which also has the greatest cone load, can be carried to considerable distances by even relatively light winds.
Normally, Phymatodes does not eat the seeds although it may damage some of them in its feeding pathway. As mentioned earlier, browned cones do not necessarily drop their seeds immediately upon opening, but rather drop them slowly over the years as the vascular connections are severed.
The beetle larvae average 1.4 individuals per cone in the cones attacked, and Stecker has occasionally noted as many as eight in a single cone. About one-quarter of the 39,508 cones in the Castro Tree are "browns," or cones on which Phymatodes has fed. If this is more or less representative for mature sequoias, this tiny insect deserves considerable credit for spreading the sequoia's seeds. It seems that each species' well-being depends on some reciprocal arrangement of services.
Chickarees, interestingly, seem to prefer cone flesh when the cones are between 2 and 5 years old, infrequently cutting 1-year cones, but for some reason seldom eating them. As Phymatodes is apparently most prevalent in cones 4 years and older, there is really little competition between these two animals. This further insures that seeds of all age classes are shed. The sequoia, the chickaree, and the cone beetle may someday reveal a fascinating coevolutional story because without the two animal organisms, as well as frequent fires, the giant sequoia might not exist today.
We know that the chickaree prefers the larger seeds of other conifers, and that it feeds more heavily upon the cone flesh of sequoias in years when the other food is in poor supply. If a fire destroys a large percentage of the other tree species, chickarees provide added assurance by eating sequoia cones and dispersing the seeds at a time when the seed bed is a soft, friable mineral soil. Fires also speed the rate at which cones will dry out and scatter their seeds to the ground. In essence, this is a back-up mechanism insuring seed release and rather heavy seeding of sequoias after a fire.
While animal activities cause most of the seed dispersal, cones do fall during wind and ice storms and because of heavy loads of wet snow. The proportion of this kind of seed-fall has never been measured. Certainly it is locally heavy, but occurs so irregularly that its overall role in regeneration may be slight in comparison to animal-influenced release. Some cones find their way into streams and are carried considerable distances, extending the range of the species. (See details in sections discussing present distribution.)
What happens to a cone and its seeds if not cut by a chickaree or opened by the feeding of Phymatodes is a minor question. Note that, although the oldest known cone in a green, growing condition was 22 years of age, the longer the cones persist on the tree, the less viable their seeds; hence, increased length of cone retention does not benefit the species. Furthermore, the older cones frequently become so encrusted with foliose lichens that the seeds are less able to fall free, even when the cones finally open.
Seed viability loss after dissemination
Whereas seed viability in green mature cones may be as hight as 75%, snap tests of seeds taken from the surface of the soil indicate an average viability of between 1 and 2%! This markedly reduced reproductive potential seems due mostly to the sun's direct radiation and to desiccation resulting when radiant energy is converted into heat at the soil surface. When kept dry and out of sunlight, sequoia seeds are known to maintain their viability for many years.
In tests, a viability of 40% dropped to 10% when the seeds were exposed to sunlight for 10 consecutive days, and 0% by the end of 20 days. Although this reduction in viability is proportionate to the size of the seeds, the average sampling of seeds tested shows clearly how vulnerable is this stage of the sequoia's life cycle. Thus man, by changing the tree's environment, can either aid or retard its reproduction rate.
Effect of animals on seeds upon the ground
Contrary to some earlier reports, animals do not greatly disturb sequoia seeds on the ground. Insect damage to the embryo is probably the most serious effect noted to date, but we do not know whether this occurs before or after loss of viability by radiation and desiccation. Vertebrate animals apparently rarely eat the seeds on the ground. Repeated tests by Howard Shellhammer and by Beetham (1962) both in and out of sequoia groves have shown that animals favor sequoia seeds least among the seeds of sugar pine, ponderosa pine, Jeffrey pine, red fir, white fir, and incense-cedar. Usually, if sequoia seeds are placed on the ground in piles, squirrels, chipmunks, mice, or shrews merely kick them about and leave them uneaten.
Conditions for germination
For germination to occur, the giant sequoia requires the necessary physical conditions, but also the proper sequence of events. Perhaps even more uncertain is the survival of the resulting seedlings.
Muir (1878) was very probably the original source of the idea that bare mineral soil is essential for the germination of sequoia seeds. This was a convincing story, which virtually all sequoia literature repeats without question. Yet, in some sequoia groves, there are extensive areas of almost bare ground within the seeding range of mature sequoias on which we find no sequoia seedlings year after year. In other instances, mineral soil plays absolutely no role in germination and small sequoia seedlings growing from rotting stumps and other masses of organic debris such as thick leaf litter. Normally, such debris holds too little moisture to permit continued growth; if the litter is too thick, the seeds' small energy package does not allow the seed roots to penetrate to mineral soil and, if germination does take place, the young trees often die aborning.
Mineral soil, then, is only one of several influences upon germination of sequoia seeds. Statistically, it is the most important substrate for germination, but is not an absolute requirement. The condition of the bare mineral soil, also affecting the survival of seedlings which do actually get started, may well be at least as critical, if not more so, for seedling survival as it is for germination.
To better insure germination, the soil's condition must be such that some sort of disturbance will loosen it before the distribution of seeds. The tiny seeds lack sufficient weight to become buried upon falling, except where the soil is very soft and friable. This perhaps explains the absence of seedlings on soil surfaces where the seeds may be unable to penetrate. When lying on the exposed soil surface, the seeds not only quickly lose their viability but also seem to germinate poorly even when the soil is moist. Fry and White (1930) claim that seeds pressed against the soil surface by heavy snow germinate well, but recent experimentation indicates that germination is greatest when the seed is completely surrounded by moist soil, as when burried. That seeds germinate on the surface is well documented; but again, survival is extremely low.
Disturbances of the soil may come about in any of several ways. Falling trees leave exposed and loosened soil in their root pits, and the skidding trunks on sloping terrain may "plow" up the soil and even bury some seeds. Avalanches of snow and floods of water contribute to the burial which favors germination. But fire is perhaps the most influential and certainly the most widespread of all the natural factors. In fact, there is a high probability that, without fire, the giant sequoia would not today be an extant species.
Fire burns the organic content of the uppermost horizon of the soil leaving temporary voids between the soil's particles, so that the tiny seed falling from the crown is often buried to a depth sufficient to insure its full contact with the soil and to eliminate potential radiation damage. Usually, the soft, friable surface condition of the soil after fire is short-lived; rainfall, wind, and gravitational settling of the soil particles result in compaction, which in turn reduces successful germination. We have also observed that the heat of fires speeds the cone's drying and that seed-fall during the days immediately following fires is greatly increased.
In recent years, of course, man's mechanical disturbances to the soil—as along roadsides, in areas of building construction and logging activity—have likewise created a receptive medium for seed burial, so that seedling growth is often extremely heavy and the degree of survival is high.
Other factors that influence germination, for better or for worse are air temperature, soil moisture, light, mineral content of the soil, soil pH, soil type (texture), and depth of the seed in the soil. These, and other lesser influences, have been thoroughly investigated by Stark (1968) in the Sierra Nevada.
Stark's field experiments indicate that although germination actually occurs over a very wide range of temperatures, namely, from 30° to 92°F (-1.6°C to 34°C), optimum temperatures are most common during the months of April, May, September, and October. Soil moisture conditions and seedling survival are generally better in the spring than during any other season. High summer temperatures and the resulting desiccation of the soil greatly reduce germination.
Soil moisture content varies with soil texture and the amount and nature of the soil's organic matter. At field capacity, which is the maximum moisture that can be held against the pull of gravity, native Sierran soils, which are mostly sandy, probably store no more than 18-22% soil moisture by weight. This appears wholly adequate for the germination of sequoia seeds, and the high sand content insures good aeration in the soil, which is another requisite for successful germination. In flooded soils there is little oxygen available for the embryo's respiration and growth. Some seeding occurs along streams, but germination occurs only when floods deposit seeds high enough on the banks to escape the flooded soil conditions. Dry soils also reduce germination to the zero point, although the critical minimum level of moisture for this process is unknown.
Experiments indicate that sequoia seeds will germinate in full sunlight and also in the dark, but that optimal germination occurs during the growing season when the light is approximately one-half full strength of the sun. When stronger, light is converted into excessive heat energy and thus dries the soil. Again, fire plays an important role in preparing the seedbed by reducing the amount of shade. Under field conditions, sequoia seeds require between 40 and 60 days to germinate.
Whether natural chemical inhibitors affect the germination of sequoia seeds on the ground is not yet known. In preliminary studies at San Jose State University, germination tests subjected sequoia seeds to various solutions of chemicals normally found in soils and to solutions extracted from leaf litter (Morris 1967). There were no significant differences over those in the controls, indicating possibly that, in the natural state, physical factors influence germination more than chemical factors. Closely allied is the pH or degree of acidity or basicity of the soil, Stark (1968) found that a slightly acid soil (pH 6-7) produced the highest germination percentage at a temperature of 68°F and concluded that pH was not a limiting factor in natural sequoia habitats. She found that strongly basic soils (pH 9) stunted the seedling growth, but did not retard germination. It did, however, alter the color of the foliage to an intense blue-green.
Variations in soil texture are actually not very important in the sequoia's native Sierra Nevada where sand percentages are high and clay percentages characteristically low, a combination admirably suited to successful germination. Experiments of Beetham (1962) indicate that poor germination in clays, limestone, and peat soils indicates relationship to insufficient moisture. Once established, seedlings transplanted to many other soil conditions grow well throughout the world. Sequoias have never re-seeded themselves naturally on these other soils, indicating this tree's special needs for successful reproduction.
We have already mentioned that seeds on the surface generally do not germinate because insufficient moisture is transmitted to the embryo. Burial of the seed is important, but the seed must not be buried too deeply. While seeds placed deeper than 1 inch may germinate, the developing shoot will seldom reach the surface and survive. The optimum depth, which seeds rarely exceed in normal circumstances, is about 0.25 inch (Beetham 1962).
Seeds will often become wedged in a small crack in the soil alongside a partially buried rock or piece of wood, which provides the necessary protection against radiation and proper soil moisture conditions. This is also an advantage in seedling survival, the next most delicate stage in the sequoia life cycle (Hartesveldt and Harvey 1967).
Conditions for seedling survival
. . . fire, the great destroyer of the sequoia, also furnishes the bare ground required for its growth from the seed. (Muir 1878, 1912).
To a very strong degree in sequoia reproduction, seedling survival is much more critical than is germination (Beetham 1962). Muir alluded to this in 1878, and numerous writers since have repeated it. Sequoia seeds can remain viable under a rather wide range of conditions, but once the seedlings commence their growth, they cannot survive beyond the range of a rigid set of environmental conditions. It is now suspected that the microenvironmental climate is the major restriction squeezing the species into the present isolated groves, and that it has limited the groves' expansion to relatively minor boundary extensions during the last century or so. Some believe that sequoias would doubtless flourish in many locations in the Sierra Nevada outside the existing groves if stock were only introduced. According to this hypothesis, extension of the range is limited by a subtle environmental barrier and one that was poorly understood until the recent outstanding work of Rundel (1969)
Because of the small amount of food stored in sequoia seeds, the newly germinated seedlings must become rapidly self-sufficient. Fry and White (1930) state that the earliest stage of germination (extension of the radicle or primary root) takes place beneath the snow, and that the seed roots are as much as 1 or 2 inches long before the snow melts. This may affect the survival of the emerging cotyledons, or seed leaves. As soon as the protective seed coat is shed from the new leafy shoot, a root system must be functional to supply the cotyledons with the necessities for photosynthetic activity, and because sequoias apparently produce few, if any, root hairs, root length becomes the more essential.
Considering the many hazards offsetting this species' great reproductive potential, we may surmise that survival of seedlings is tenuous. Muir (1878) records that not one seed in a million germinates, and that not one seedling in 10,000 attains maturity. These figures, widely repeated, may be figurative rather than literal, but the frailty of the species during this stage is no myth.
The newly germinated seedlings are, like those of all plants, tender and soft because of the yet small deposits of cellulose and lignin and are susceptible to a variety of decimating factors even before the unfurling crown is pushed above the soil surface. Some seedlings seem more predisposed to survival than others. For those that die, the cause of death is not always obvious, especially if the roots are affected. Furthermore, the decimating factors may be subtly and confusingly interrelated. Seedlings derived from the larger seeds may have the advantage of a larger and more rapidly growing root system from the very beginning and, therefore, a better chance for survival. However, our studies indicate that even the heartiest of seedlings may die in areas where others of apparent lesser vigor will survive.
Probably the most extensive cause of sequoia seedling death is soil desiccation downward to the full depth of the root system. Harvey suggests other possible causes are damping-off, intolerance to shade, flooding, heat canker, root fungi, soil ectocrines, burial by leaf- and branch-fall, insect depredations, grey mould blight, being eaten by birds and mammals, trampling, and various other disturbances by animal life, including man. Many of these are probably more serious threats where there is deep duff and litter. Fire, on the other hand, in removing the duff and litter reduces some of the danger.
Desiccation of the soil is generally more prevalent in disturbed open forests, e.g., following a fire. It is related to the length of time the sun strikes the mineral soil surface during the day, to air temperatures, relative humidity of the air, and to some extent the color of the soil surface. Fire has been the most common natural agent influencing the above factors and, in some cases, it also darkens the surface with its char. The increase in sunlight, so necessary to the survival of sequoia seedlings, is also the factor which may dry the soil to the permanent wilting point and bring about the seedlings' death. And while the litter layer is an insulation against soil moisture depletion, it undoubtedly harbors the damping-off fungi and other pathogens (Martin 1957-58). Obviously, sequoia reproductive success lies somewhere between the two extremes. But nature is generally not much given to continuous provision of optimal conditions for survival of any one species, and where sequoias fail, other plants with adaptations more suitable for the immediate set of conditions may succeed.
Beetham (1962) has amply demonstrated that seedlings grow best in full sunlight where the soil is protected by at least a light layer of leaf litter. Where litter was absent, she found soil temperatures 25-35°F; hotter and increased death rate of seedlings by heat canker. Hartesveldt et al. (1967) recorded surface temperatures in July up to 157°F on char-darkened soil in the Redwood Mountain Grove at 1:45 p.m. by means of a tiny thermister, which records the temperatures of a literally paper-thin layer at the soil surface. The threshold temperature for the death of most protoplasm being less than 150°F, not surprisingly, several seedlings were found dead with the blanched and sunken symptoms of heat canker on the stem just above the mineral soil level. Despite the very high temperature of the soil surface in this open situation, many seedlings in the same vicinity were not killed by the heat, a testimony to litter effectively moderating temperature extremes.
By far the greatest mortality occurs where soils dry out to below the seedlings' average rooting depth, which is rather common during periods of prolonged though not necessarily excessive high temperatures and low relative humidity. In experimental manipulations, Hartesveldt et al. (1967) found that more than 90% of the seedling mortality occurred under these conditions. Death began within a few weeks after germination, continuing at a much reduced rate in the following years. At the end of 3 years, surviving seedlings will usually have root systems that penetrate the soil to beneath the level of midsummer dryness, or about 14 inches.
Beetham (1962) reported that optimal growth of sequoia seedlings occurred in soils with moisture contents at or near field capacity, or about 20% for most Sierran soils. She further determined that the soil moisture content at the time of seedling death by desiccation is about 5.2%. Seedling survival was found to be critical in the low range of soil moisture in studies of Hartesveldt et al. (1967) in which desiccation was the most frequent form of seedling death.
However, those same studies revealed that relatively slight variations in soil moisture in the lower range may make the difference between survival and death. It was found that seedlings growing next to partially buried rocks, limbs, etc., definitely had a greater advantage in being taller, more branched, and surviving better than their nearby counterparts whose stems were surrounded only by soil. This may be a response to better soil moisture conditions beneath the partially buried objects, perhaps due to the deflection of drying winds and the interference of direct solar radiation at the soil surface. Together, these factors reduce the rate of evaporative moisture loss from that portion of the soil surface where the objects occur.
We should point out that Sierran soils are notably poor compared with those measuring up to agricultural standards, and limitations to seedling growth apparently do not relate to low nutritional levels in the field (Beetham 1962).
One of the more difficult forms of seedling death to assess is that from reduced light brought about by canopy shading that may starve the plant. Although Baker (1949) lists sequoia as having intermediate tolerance to shade, Beetham (1962) indicates clearly that it is very sensitive to low light intensity. This is supported by the fact that sequoia seedlings are seldom found in areas densely populated with taller vegetation. A striking example of death influenced by shading was found by Hartesveldt (1963) in Yosemite's Mariposa Grove. At the end of a 25-year period, of the several thousand seedlings established there and recorded on a park map dated 1934, only 13.8% remained alive in a 1959 resurvey of the same area. Hundreds of dead saplings, twisted and contorted in dense shade, demonstrated the effect of the heavy overtopping crown canopy composed largely of white fir (Fig. 10). Soil moisture appeared adequate in the areas in which the young sequoias had died. Undoubtedly, excessive shading may be coupled with another agent such as root fungi or poor soil-moisture conditions. In marginal circumstances, young specimens barely maintain life and grow so sparingly that measurement of yearly increment is difficult. In fact, Metcalf (1948) records a 25-year old specimen as having a stem just 0.50 inch in diameter.
Excessive moisture is a factor which limits gas exchange at the root surface because it usurps the pore space normally occupied by gases. Low soil oxygen content reduces root respiration which reduces water intake and photosynthesis, eventually to the point of cessation. This is probably a common cause of seedling death along the edges of meadows where seeds of sequoias are often scattered abundantly, but where seedlings seldom survive. Although the situation has not been thoroughly studied, the general lack of trees within the wetter meadows is expressive of the sensitivity of most tree species to wet soil. Moreover, dead sequoia snags are occasionally found in wet meadows in which the meadows have formed after the tree had become established in a more mesic situation. Large sequoias often fall across drainageways, forming a dam which impedes water drainage and creates wet meadows. Meinecke (1927) recorded the death of large sequoia trees by this means in the Giant Forest.
Where sequoias do become established in moist areas, the degree of moisture definitely affects their growth, and even large specimens have died where the soil moisture has become excessive.
Soil dampness promotes the incidence of damping-off, a disease which fells the seedlings by attacking the stem at the soil level. Any one of several soil fungi can be causative agents in this disease, Damping-off has long existed in forest nurseries among virtually all types of trees, and the giant sequoia is no exception. Another disease affecting juvenile sequoias is a root rot caused by the fungus Sclerotium bataticola Taub. found in damp, dark areas. Gray mould blight was recorded by Martin (1957-58) on sequoia seedlings in Germany, and we later observed it in the Giant Forest and in the Redwood Mountain Grove. The affected leaves appear "cemented" together by the fungus, which completely destroys their photosynthetic capabilities. Long burial under wet snow in winter and spring is the probable cause. For overall seedling survival, however, it is only a minor problem.
The depredations of insects and vertebrates on sequoia seedlings have been exaggerated because of the statements by Fry and White (1930), which we have not been able to verify. Their observations in the park nursery at Ash Mountain, outside the tree's natural habitat, possibly led to the contention that no other conifer is attacked in its infancy by so many destructive agencies. They further state:
Birds, such as finches and sparrows, are attracted to the green tops for food. Cutworms destroy many of the plants by cutting them off near the ground. Large black wood ants levy a heavy toll by cutting off the plants and dragging them to their nests. Rodents, such as ground squirrels and chipmunks, destroy many of the plants by eating off their tender tops.
Insect depredations on seedlings do occur, but they appear minor and are almost wholly limited to first-year seedlings' tender tissues. Damage may also be accentuated when sequoia seedlings predominate among foods available, as they may after a fire. In weekly post-manipulation examination of hundreds of dead seedlings, from 3.4 to 17.5% per year showed signs of insect feeding. The tender epidermis on the stem was commonly eaten away from the crown downward to the ground level, or else the stem was girdled. In a report of ours (Hartesveldt et al. 1968), Stecker identified this damage as the work of a camel cricket (Pristocauthophilus pacificus Thomas), a nocturnal feeder, in its first and second instar stages. Occasionally, the larvae of a geometrid moth (Sabulodes caberata Gn. and Pero behrensarius Pack.) were feeding upon the seedlings' leaves, but the few resultant deaths would qualify it as relatively unimportant.
There is some indication that tops of new seedlings are eaten off by birds or mammals, but the number is certainly insignificant, as is the number of those uprooted by rodents or by deer hooves. Compared to desiccation losses, animal losses are negligible.
Sequoia seedlings probably seldom die by winter freezing within their native range, Temperatures here rarely reach 0°F and, at times when such lows are most likely, seedlings are generally well insulated by snow and are not much affected. Beetham (1962) lost seedlings planted above 9600 ft in the Sierra where low winter temperatures prevail. Low temperatures are more a problem in nurseries (Wulff et al. 1911) and where seedlings have been introduced into cold climates in other parts of the world.
Seedling density
The density and rates of growth of young sequoias vary considerably with the circumstances of seed distribution and seedling survival, Where fire has created optimal conditions for seedling establishment, there may be as many as 25-50 seedlings, and reportedly even more, per square foot. Such densities, however, are usually limited to relatively small areas because of either an irregular seed dispersal pattern, or spotty soil receptivity, or both. In areas where fires have heated the soil surface, the high temperatures seem to favor both the seedlings survival and increased rates of growth.
DeBano and Krammes (1966) have discussed water-repellent soils and their relationship to fire; more specifically, Donaghey (1969), experimenting with soils from sequoia groves, has demonstrated that incineration of soils increases both their wetabiity and soil moisture retention. This better explains sequoia seedlings' survival and growth in burned areas. On recent experimental and prescription burns in Kings Canyon National Park, seedlings have been especially abundant in soils severely burned by the combustion of dry downed logs, and their resulting elongated pattern conforms to the position of the log (Fig. 36). This high seedling survival under burned-out logs possibly explains the remarkably straight rows of even some mature sequoia trees.

|
| Fig. 36. Counting seedlings in trough of burned sugar pine log. The seedlings are too small to be evident in the photograph, but author Ron Stecker and wife Phylis recorded young seedlings in the soil heated by its combustion. Records indicate that seedling survival is better in situations where soils have been heated to a high temperature. This could well be one of the reasons behind the straight lines of mature sequoias. |
Competition, of course, increases with density, and mortality thins out the trees continuously as their crowns expand and compete for light, Being very intolerant of shade, the young trees without sufficient light are killed, or at least the lower portions of their foliage die away.
The juvenile stage is commonly a narrow conical spire where site conditions are optimal and where growth is therefore rapid (Fig. 11). When sequoias are virtually the sole species established in a given area, the slender crowns are well-fitted for maintaining dense stands for many years since the conical form assures that the sun will reach part of each crown. The 10-year olds portrayed in Fig, 37 at Cherry Gap, Sequoia National Forest, are from 6 to 15 ft tall, and such stands are much too dense for a man to walk through with comfort. Although the supply of subsurface drainage water is highly dependable, this density obviously cannot long continue because the light factor will soon become critical for the smaller trees. Those which now dominate the others in height will almost assuredly be the eventual survivors. Of the 126 saplings in this small grouping, probably no more than four or five will survive to mature size.

|
| Fig. 37. Sequoia reproduction in area burned by fire of 1955 in Cherry Gap area of Converse Basin. Of the 126 saplings in this small group, few will survive. Photo by H. Thomas Harvey. |
The sapling stage
In areas of high site quality, vertical growth of young sequoias is commonly as much as 1-2 ft per year, and rapid growth continues until the trees are 100 or more feet high.
The trunk grows rapidly in thickness at the same time, the growth layers of vigorous specimens being as much as 0.50 inch thick or more. Both radial growth and growth in height reflect photosynthetic success. Radial growth decreases slowly with size and age although total additions of wood, spreading over a greater circumference, may remain essentially the same.
At this stage, cone production is usually well advanced and the vigorous spire-topped trees may be heavily laden with cones of several years' production.
As the spreading bases of the crowns begin to compete for light, not only does the foliage begin to die back, but within several years' time the limbs are self-pruned through shading and give the tree a more mature appearance, In its native habitat, topographic and soil conditions rarely restrict nearby competing vegetation to such an extent that a sequoia maintains its branches down to the ground level. Specimen trees in municipal parks and other open situations show this characteristic much more commonly than in their native range (Fig. 30).
Maturity
As vertical growth slows, elongation of the lateral limbs continues at an increased proportional rate so that the crown's sharp spire gradually gives way to the rounded form of the mature tree (Fig, 16). If the tree is more than 200 ft high, it has, in all likelihood, solved its light problem and, barring the catastrophies of intense fire, lightning, snow-loading on the crown, and blow-down, now stands an excellent chance of surviving to old age. No other tree in its native Sierra Nevada overtops the crown of the taller sequoias, Thus, its upper foliage, which is still intolerant of shade, has a dependable, uninterrupted supply of sunlight for its photosynthetic needs.
By this stage in the sequoian life cycle, the total number of large, mature specimens has been considerably reduced by fires and other decimating factors. In the Redwood Mountain grove we studied, the mature trees varied from 5 to 8 per acre. Elsewhere, in localized situations, the density may approximate 15-20 per acre such as the Senate Group in the Giant Forest and the Sugar Bowl group in the Redwood Mountain Grove. Nowhere has the sequoia ever been found as a pure stand.
Old age
The characteristic form of the older trees is one of irregular, craggy, and sometimes grotesque crowns which often reveal the dead upper part of the trunk, or "snag-top" (Fig. 38).

|
| Fig. 38. "Snag-top" sequoia, indicative of old age and injuries to the base of the trunk. Older sequoias have felt the ravages of fires over and over again. Where burn scars interrupt the flow of soil nutrients, often the uppermost portion of the tree dies back. Photo by R. J. Hartesveldt. |
Although lightning has been suggested as the reason for these dead tops, it now seems more probable that they are the result of fire scars at the bases of the trees which have a diminished translocation of water upward from their roots. The few large, old specimens still maintaining symmetrically rounded crowns invariably have small fire evidence, usually only a superficial char on the bark. A study of 100 snag-top sequoias in the Giant Forest revealed that each of them had one or more basal fire scars burned through the bark into the wood tissue (Hartesveldt 1965). The severance of the effective connections between the root system and the crown may well have deprived the crown of a portion of its requisite supply of moisture and nutrients (Rundel (1971), and it seems logical that the portion with the longest supply route would be the first to suffer. Scars at the bases of snag-tops are from 15 to 96% of living specimens' total circumference and there is a general correlation between the size of the scar and the size of the dead snag at the top.
It is very probably impossible to find a sequoia in the mature or later stages that does not bear the black char marks of fire. Various authors have strongly suggested that fire has been a normal factor in almost all Sierran environments since the time forests existed there. The degree to which a tree is burned or spared probably depends on its topographic location with respect to other vegetation, the relationship to fuel accumulations, and other factors which would similarly influence the intensity of the often-repeated fires. The burn scars of individual trees are probably due more often to the cumulative effects of many fires, not all necessarily intense, than to single fires.
Prior to the Western innovation of fire prevention and suppression, both the incidence and size of forest fires were greater than now. The intensity, however, of fires then was probably less. With repetitions of fires at rather regular intervals, fuel could not accumulate to proportions that would support today's dreaded crown fire holocausts. Each fire cleaned up the previous accumulations of fuel on the ground, and although other trees' crowns may have been badly damaged, sequoias, because of their size and fire-resistant bark, had a substantial survival advantage. Individual, standing, dead sequoia snags are common and show clear evidence of having been burned to death. But such dead snags are rare in a given locality, and nowhere cover a large area, suggesting that intense crown fires were less frequent in the past.
In the snag-top study referred to earlier, fire scars were found just about in any conceivable location on the trunks observed. They varied from surface scorching to deep "cat-face" scars only at the base to longitudinal scars often running the full length of the trunk, and to hollowed "telescope" trees. Of the basal scars, analysis showed that approximately 90% were decidedly on the trunk's up-hill side. In the field, the reason becomes quite obvious. Cones, limbs, and other combustible debris, as it falls to the ground, is generally carried downhill by the pull of gravity and it comes to rest against the barriers formed by the trees' trunks. Thus, even light ground fires smolder longer in these collections of fuel and sear slowly through the bark and into the wood tissues.
The resulting scars on the up-hill side become somewhat concave just above the ground level and the cavity presents a greater storage space for new accumulations of falling fuels. Therefore, not only do later fires have a greater fuel supply, but the concave surfaces also tend to accentuate the effects of subsequent fires as they reflect the heat like an oven. And the larger the scar, the more pronounced the effects of each successive fire.
Despite their cragginess and "imperfections" in old age, the trees remain mystically beautiful and enchanting. They are by now the largest members of their community and they dominate the tree-top horizon in such a manner that the species can be identified easily by an observer many miles away. These are the specimens of trees that prompted the establishment of the parks that now preserve them and are the trees which are most prominent in the interest of the visiting public.
Contrary to popular belief, these old veterans have not stopped growing, nor have they necessarily even slowed their rate of growth. Green foliage proclaims that photosynthesis is still on-going, and this, in turn, implies continuing yearly increments to the trunk diameter, no matter how large the specimen. Although the annual ring width is narrower, the radius of the tree becomes continually greater so that the amount of wood added each year is proportionately the same as when the tree was younger and is more or less maintained continually.
The ring width at this stage may vary from 0.50 mm per year to 1 mm for trees in well-watered sites. This means that during each 24 years the world's two largest trees, the General Sherman and the General Grant, will add about 24 mm (1 inch) to their radial growth, or 48 mm (2 inches) of diameter. Even the fragmental trunk of the Black Chamber in the Giant Forest is adding 1 mm of radial growth yearly, despite 96% of its circumference having been destroyed by fire, as is most of the trunk (Fig. 39). Here is the quintessence of tenacity of life for which this species is so well known.

|
| Fig. 39. The Black Chamber, Giant Forest, Sequoia National Park. This tree is a living remnant of its former 18-ft diameter, with 96% of its former circumference burned away. Yet the tree is growing radially at about the same rate as an uninjured tree of similar size in optimum site condition. |
Unlike other plants and animals, the sequoia does not seem to lose its reproductive ability with old age. The very largest specimens (not necessarily the oldest) have crowns that are loaded with cones containing viable seeds. Difficult as it is to determine the age of living trees without considerable effort, it is likewise difficult to assess any slowing of cone production due to aging. But, over our years of observation, we have found no indication of such slowing. The five largest specimens of sequoias in the world all bear moderate to heavy cone-loads and germinable seeds. Likewise, the remnant crown of that hopelessly "crippled" Black Chamber is loaded with green cones.
That lightning affects the tops of tall sequoia trees is well documented by observers fortunate enough, from our point of view at least, to have been near one of these natural lightning rods when it was struck. Fry and White (1930) describe a dramatic horizontal bolt of lightning striking a trunk with a 16-ft diameter in the Garfield Grove. Surely it was the sight of a lifetime, the bolt knocking out a 20-ft segment from the middle of the trunk and dropping the crown which, oddly, caught up in the split of the trunk below. They further report that the trunk sent out new growth and may be presumed alive today. The Giant Forest seems to have a higher incidence of lightning strikes on sequoias than most of the other groves, perhaps because it is so close to the edge of the great "plateau" on which it is located. In several trees broken off near their middles, the lower branches have taken over the crown's sole function (Fig. 40). Recovery for these stricken trees appears reasonably well assured.

|
| Fig. 40. Lightning-topped sequoia, Giant Forest, Sequoia National Park. In contrast to the "snag-topped" sequoias, those struck by lightning have had at least the upper third of the trunk broken away. Lightning seldom kills mature sequoias in the Sierra Nevada. Photo by R. J. Hartesveldt. |
Former National Park Service employee Ralph Anderson recalls vividly one of his earlier days in the service when he was stationed in Yosemite's Mariposa Grove. During a summer thunderstorm, a bolt of lightning struck the dead top of the Grizzly Giant, dislodging several hundred pounds of wood and dropping them literally at his feet. Although this incident may have strengthened the assumption that snag-tops are created by this means, it must be recognized that where bolts of lightning have hit the living parts of trees, the entire upper portion was often broken away, perhaps because of the great heat affecting the moisture within the stem. The Grizzly Giant's dry top, dead many years or even centuries, was perhaps a poor conductor of electricity, resulting in a relatively small portion of wood being torn loose. Fry and White (1930), perhaps the longest continuous observers of the giant sequoia, record only two specimens actually killed by lightning.
Differing from most other tree species, the sequoia seems to have no known age of senescence. There is no record of mature sequoias ever having died from disease or insect depredations, afflictions common to other species of trees in their old age. If the sequoia could be kept from falling over or being burned to death, we can imagine that our descendants, a millennium hence, may see a General Sherman Tree approaching 40 ft dbh.
Death
With advancing old age, the thick, resin-free bark of this tree is doubtless a definite asset to its longevity and survival. Fires that kill or severely damage other species and younger sequoias may be of far less consequence to the relict larger sequoias which have survived many successional sequences of these associated species. Occasionally, however, a fire has been severe enough to kill all the foliage on more than one specimen in a given locality. Stricken, charred trunks remain witness of these fires for many centuries before they disintegrate. Seldom are more than two or three such snags found in close proximity, which suggests that crown fires were less prevalent before the advent of Western Civilization.
Sometimes lightning strikes will set fire to the dead wood of a snag-top specimen. The wood, which may have been drying for a thousand warm Sierran summers, holds the fire very well, contrary to popular opinion and despite the limbs' size. We watched such a fire in the top of a large sequoia in Redwood Canyon in the summer of 1966. Charles Castro of the Sequoia National Park Forestry Division had climbed into its crown via a nearby fir tree to spray water onto the burning portion, unreachable from the ground. From our vantage point, and with Castro as a reference, the burning limb seemed about 3 ft in diameter, hardly a good prospect for a continued blaze. Just such a fire in the Giant Forest Lodge cabin area burned away the entire crown and upper part of another large sequoia in 1959. Again, the fire was burning well above the reach of water streams, and there were no adjacent trees suitable for access into its crown even if Castro had already developed his unique fire-fighting method. The tree, which burned for 2 weeks and disconcertingly dropped firebrands amongst the cabins, is now but a dead snag to about one-third its original height.
Old sequoias most commonly die by toppling. Because of the wood's brittleness, this virtually assures death, although an occasional specimen has retained green leaves for several years after falling, which perhaps indicates that the fall did not sever all its vascular connections. One unique tree in the Atwell Grove that fell many years ago is still growing vigorously, with seven lateral branches giving the appearance of a candelabrum.
Fire is probably the greatest contributor to death by falling because it creates gaping fire scars at ground level which weaken the tree's mechanical support. The extreme weight of the big trees coupled with their shallow roots increase the effects of this weakening, especially in leaning trees. Other causative factors are water-softened soils, undercutting by streams, snow-load on the crown, uneven reloading with moisture, wind, heart-rot, nearby falling trees, perhaps carpenter ants cutting galleries in the bark and dead wood near the trunks' bases, and, of course, various combinations of the above.
We mentioned the finding that 90% of the fire scars in trees growing on slopes were on their up-slope sides. It is not surprising, then, that 90% of all fallen trees seen during that survey were found to have fallen up-hill, toward the side with the least support. Likewise, sequoias on the edge of wet meadows tend strongly to fall into the meadow because of the greater weakness associated with the softened saturated soil, and, in addition, because of the heavier foliage on the more illuminated side. Trees in these soft substrates also may come to lean because of wind pressure or snow-loading on the crown. If they survive, their off-center of gravity still continually subjects them to ever greater strains. Yet many of them, such as the immense Grizzly Giant leaning about 17° from the vertical, marvelously remain standing for centuries. The large, tenacious roots obviously provide sufficient anchorage to prevent their fall.
The lean of the famous Tunnel Tree in Yosemite National Park was surely part of its undoing, to say nothing of the tunnel cut through it in 1881 which weakened the tree much as a fire scar would. Unfortunately, this tree was chosen because of its large fire scar which lessened the work required to carve the tunnel. The Washburn Brothers were paid only $75 for the task (Russell 1957). Less wood had to be removed on the up-hill side, and the tree had a decided lean in that same direction.
Examination of its remains revealed a failure of the wood on the leaning side, which was also the one with the least wood support. The tree probably collapsed between February and May 1969 when, very fortunately, park visitors were not lined up bumper to bumper awaiting their turn to park in the tunnel and take the traditional photograph to be found in almost all geography books for nearly a century (Fig. 41). Because of the unusually heavy snow during that winter, the crown may well have borne 1-2 tons of additional weight which the wood could not support. No evidence supports the idea that its collapse was due to excessive trampling by people, the possible effects of which had earlier caused considerable concern (Hartesveldt 1963). Of course, the cutting of the tunnel is looked upon today as a sort of vandalism, and the tree very likely would still be standing had the tunnel never been cut.

|
| Fig. 41. The remains of the Wawona Tunnel Tree following its collapse in 1969. After surviving 88 years of traffic passing through its well-known tunnel, it was felled by the rigors of winter. Photo by R. J. Hartesveldt. |
Curiously, although averse to vandalism to our irreplaceable natural objects, many people held the Tunnel Tree in near reverence. In both Yosemite and Sequoia National parks, visitors have most often asked, "Where is the tree I can drive through?" Despite this question's humorous association for rangers in Sequoia National Park, the announcement of its falling was greeted with great sorrow.
Its fall marked the end of an era in Yosemite that many will remember fondly. Letters have often inquired when and where the next "tunnel tree" would be cut. Of course, it will not be, at least not in the national parks and forests or other public lands because laws hold federal agencies responsible for the preservation of these near-immortal trees. Not far north of the Grizzly Giant in Yosemite's Mariposa Grove is the tunneled California Tree, which in many older photographs bears the name "Wawona" over its portal. This was a useful ruse perpetrated by stage drivers in the spring of the year when the upper part of the grove was still deep in snow and the real Wawona Tree was inaccessible. Delighted passengers failed to notice that the two trees were really quite different. To assure better preservation, this tree has long been accessible only by foot.
One rather frequent form of toppling difficult to explain occurs during the warm summertime when the air is still. Records on individual trees are few, and so cannot tell us whether all or most of the fallen trees were leaning ones. If they are leaning, why haven't they gone down at some earlier time? Just what finally gives gravity the advantage after many centuries and brings the trees down? The late Willis Wagener, formerly of the U.S. Forest and Range Experiment Station at Berkeley, proposed some initial hypotheses and submitted them to us a few weeks before his death. To follow up these hypotheses, projected studies in the Giant Forest will consider leaning trees, excessive loss of water by transpiration on warm, dry days, and the shock of recharge weights as more humid conditions return. We hope these studies will bring determinations of and predictions for "accident-prone" specimens, and prevent such tragedies as that of August 1969 in the Hazelwood Picnic Grounds, Sequoia National Park when a woman was hit and killed in a bizarre falling of two trees on the windless day.
The chapter on the characteristics of old age may never be finished, at least for many generations to come. Continual protection and surveillance, improved age-dating for living trees, and extensive studies can determine whether there really is an age of senescence or a time when sequoias become more susceptible to debilitating diseases and insect depredations.
Man's interest does not necessarily end when a tree lies prostrate on the ground. To many, a downed tree appears even larger than when standing—witness the countless visitors scrambling over the remains in any sequoia park, and the photographs of cavalry troops, horse-drawn carriages, and automobiles standing atop the fallen logs for comparison. Long after death, these trees command admiration.
Decay
For science, the giant sequoias' slow rate of decay has raised several perplexing and still unanswered questions. The heartwood is particularly resistant to fungal attack and even the sapwood on some specimens is slow to decay. Many fallen trees appear today essentially as they did when photographed as much as a century or more ago.
The heartwood's unusually high tannin content, discouraging both fungi and insects, was first advanced as an explanation during the 1800s, and the Forest Service later gave it credence (Sudworth 1908). The hypothesis is still generally accepted (Schubert 1962). Detailed chemical analyses of substances from the coast redwood indicate that perhaps other organic substances deposited in the heartwood are also the retardants in this species (Anderson et al. 1968; Balogh and Anderson 1965, 1966). Similar studies have not been made on the giant sequoia but, considering the relationship of the two trees, we can reasonably assume that related repellents are likewise in its wood.
Perhaps the most interesting remaining question is how long the fallen trunks remain undecayed. Muir (1878) postulated that the trunks would last about 10,000 years and that their charred remains should be found in areas outside the existing groves. Such remnants, if they occur, would be a clue to the earlier distribution pattern of the species, as would the trenches in the ground made by the great impact of the trunks in falling. At this writing, no such remains to substantiate Muir's hopes of nearly a century ago have been found. Perhaps the pits or trenches have been eliminated during the milennia that erosive forces have been at work and that the log remnants would have been consumed by repeated fires. Contrary to some earlier opinions (Clark 1937; Schwarz 1904; Blick 1963), sequoia wood burns readily and individual dry logs have been observed to burn completely within a week. Furthermore, if the range were once continuous, fires must have burned repeatedly and erased every last vestige of their presence.
Some small indication of the wood's durability is indicated by carbon-dating of three wood specimens mentioned on page 56.
There is one species of tree known to be more ancient than the giant sequoia in its living state—the bristlecone pine. This five-needled pine ranges over six southwestern states, including southeastern California, although it is not found in the Sierra Nevada. Specimens growing in the White Mountain Range have been analyzed and one specimen is recorded at 4900 years of age by ring-count determination (Fritts 1969).
By cross-dating tree rings from these ancient pines and carefully accounting for missing of duplicate rings, an absolute chronology of nearly 8200 years has been assembled. This chronology has been useful in correcting the carbon-14 determinations which have apparently varied in their rates of production in the atmosphere within past ages (Renfrew 1971).
| <<< Previous | <<< Contents >>> | Next >>> |
chap5.htm
Last Updated: 06-Mar-2007