|
NATIONAL PARK SERVICE
The Giant Sequoia of the Sierra Nevada |

|
CHAPTER 6:
SEQUOIA COMMUNITY INTERRELATIONSHIPS
Structure of sequoia communities
While the interdependence of associated organisms is clear, the boundaries of what should be recognized as biotic communities are often difficult to determine and arbitrarily set.
Two major systems of community classification apply to the mountainous regions of western America. Merriam's Life Zone System recognizes that plants and animals vary in form and kind with increasing altitude, and that these assemblages are essentially in bands or zones of varying width along the mountain slopes. Originally based upon temperature zonation, this concept has limited usefulness. However, the recognition of indicator organisms for each zone may serve the fundamental concept that physical factors have selected various life forms, and that these factors, mainly temperature and precipitation, vary directly with altitude and, to a lesser extent, with aspect of the slope. Very much similar to the Life Zone concept is the plant belt system of Storer and Usinger (1963), which identifies the Sierra's plant associations in horizontal belts from the lowlands upward to its crest. Along a transect crossing such belts one would encounter the Great Valley, the Foothill, the Yellow Pine, and Boreal belts in that order.
Another system common in California is that of plant communities, such as Munz and Keck (1959) proposed and described. They recognize 29 such communities in California, each based upon the kinds of dominant plants found in association with one another. Some basic vegetation types are important here: marsh, grassland, chaparral, and coniferous forest. The latter category includes some nine recognized plant communities that range from the redwood forest in the mild coastal climate to the bristlecone pine of the much colder region of the high mountains. Under this classification, the giant sequoia is an element of the yellow pine forest community. Storer and Usinger's plant belt system places sequoias with the Yellow Pine Belt, and the Life Zone scheme, in the Transition Zone.
Because of its large bulk or biomass and tallness, the giant sequoia is clearly a dominant in its association. However, since no vascular plant species is found only in association with the giant sequoias (Rundel 1969), there is no such thing, strictly speaking, as a giant sequoia community, even though sequoia groves do qualify as biotic communities of interacting plants and animals.
The sequoia community, much like other forest communities, is stratified, or layered. The uppermost layer is the vegetation of the canopy, comprising the crowns of the giant sequoia and of other tree species such as the white fir (Abies concolor), sugar pine (Pinus lambertiana), ponderosa pine (Pinus ponderosa), and often, at lower elevations, the incense-cedar (Libocedrus decurrens). At higher elevations, it may include the red fir (Abies magnifica). In the more northerly groves, Douglas fir (Pseudotsuga menziesii) is also a part of the canopy, although uncommon in the Mariposa Grove, which is essentially that species' southern limit.
The understory vegetation includes trees of lesser stature, including the young of these species in the canopy layer, and shrubs. Of the tree species, white fir is today the most abundant of these saplings. In the earlier stages of succession, we find California black oak (Quercus kelloggii) and occasionally canyon live oak (Quercus chrysolepis). Two shrubs, perhaps the most common in the groves, that can tolerate the low light values at the lower strata of this forest are mountain whitethorn (Ceanothus cordulatus) and chinquapin (Castanopsis sempervirens) (Rundel 1969). The California hazelnut (Corylus rostrata) and a small tree, the Pacific dogwood (Cornus nuttallii), also survive well in the more shaded parts of the forest, and the northern part of the sequoia range has still another tree in the lower story, the western yew (Taxus brevifolia). The latter occurs only in the North Calaveras Grove, which is roughly the southern limit of its range.
At ground level, a wide variety of perennial plants characterizes the ground cover. Here, highly efficient photosynthesis is a necessity even though low productivity is the rule. These ground plants apparently can make maximum use of the minimal light penetrating to their level. They are rich in chlorophyll and generally have broad leaves so oriented as to expose the greatest possible surface area to the light.
The ground-cover plants vary greatly from grove to grove in kind and abundance, especially the annuals and biennials, and with the stage of plant succession. However, certain species are commonly associated with the giant sequoia in the large southern groves. The trail plant (Adenocaulon bicolor) is an associate not only of the giant sequoia but also, in the coastal mountains, of the coast redwood. Also common to many sequoia groves are sweet cicely (Osmorhiza chilensis), white hawkweed (Hieracium albiflorum), and a wood violet (Viola lobata). These four ground-cover plants are most abundant at about 5% of full sunlight in the Redwood Mountain Grove (Hartesveldt et al. 1967)—a good indication of their ability to survive on shaded forest floors. Seedlings of white fir are similarly abundant at this low light level.
In areas subjected to the physical removal of the understory or its destruction by prescription burning, two major changes occurred in the ground-cover plants. Harvey found that some of the species, such as trail plant and hawkweed, decreased considerably where the ground-level light averaged 25% of full sunlight following manipulation. Other plants increased in abundance as their light requirements became more nearly optimal.
Small-leaved ceanothus (Ceanothus parvifolius) sprang up around the perimeters of burn piles where the fire, heating the seeds, had enhanced germination. It is known from other studies (Sweeney 1956) that, in many of the chaparral species, heating the seeds changes the seed coats so that they admit water and initiate germination.
Two types of plants increased abundantly most likely because of the disturbed ground cover and exposed mineral soil. Small, delicate annuals, such as gilia (Gilia giliodes) and a popcorn flower (Cryptantha affinis), proliferated following manipulation by fire. The thin-stemmed primrose (Gayophytum nuttallii) also increased greatly. The second group, the perennials, likewise grew strikingly in number. Phacelia mutabilis and gooseberry (Ribes roezlii) showed a dramatic 10- to 20-fold increase after fires and after other physical manipulation of the ground and vegetation. Each of the above plants is part of the typical giant sequoia forest community.
The most conspicuous streamside plant within sequoia groves is the western azalea (Rhododendron occidentale). It is especially attractive in the spring, with its large, irregular, flamboyant, white flowers, and again in the fall when its foliage turns reddish. On the flood plains and gentle slopes, bracken fern (Pteridium aquilinum) and a tall lupine (Lupinus latifolius) literally carpet the forest floor. In spring and early summer, the bright purple flowers of the lupines against rustic red sequoia bark is a sight to treasure.
Within the areas studied, we find approximately 65 species of plants in the sequoia forest plant community, excluding the strictly riparian species. Although the species combination varies from grove to grove, the larger groves probably have close to that number. Appendix II contains the names of vascular plants mentioned in the text.
Plant and animal relationships
With the exception of insects and possibly some other invertebrates, animal life in the groves is very much like that found throughout the Transition and Canadian zones in the Sierra at elevations from about 4000-9000 ft. The mixed conifers support a delightful wealth of vertebrate life, much of it ubiquitous throughout the Sierra.
There is no reason to believe that any particular species of vertebrate is exclusive to the groves, even though one animal, the chickaree, depends strongly on the sequoia for food. And since no vertebrate is known to nest or roost exclusively in it, the sequoia adds little to the wildlife habitat of which it is but one constituent. The groves border on nonsequoian forest communities, as well as on wet meadows, dry rocky outcrops, open grassy meadows, dense shrubby areas, and many other types of vegetation or ecological situations depending upon local environment and the stage of plant succession.
Animal populations, then, vary with location and time according to the available food and other requisites of life provided by plants, and such a variety precludes complete and detailed analysis. What follows is a synthesis of our studies in the Redwood Mountain Grove and Whitaker's Forest, and observations which we and others made in other groves as well. It mostly describes the faunal characteristics of the typical giant sequoia grove.
Appendix III contains the scientific names of the vertebrate animals mentioned in the text.
Vertebrates
Birds are the most abundant and obvious vertebrate animals within the sequoia forests, and the composition of their populations does not vary much from north to south within the range. As Kilgore (1968) has pointed out, ". . . the bird species which are found in the surrounding forests, without either species of sequoia, make up the sequoia forest avifauna." He attributed this to bird communities' greater correlation with the growth form of the forest vegetation than with a dominant species of plant.
The most typical birds in the associated nonsequoian coniferous forest are the yellow-rumped warbler, western tanager, mountain chickadee, brown creeper, warbling vireo, Oregon junco, Steller's jay, hermit thrush, golden-crowned kinglet, and solitary vireo. Within the groves, the species composition is very much the same, but the dominance of one species over another varies with the stage of plant succession. In the young mixed conifers which include sequoia at Whitaker's Forest, Kilgore (1971) found the following four species as dominants: western robin, Steller's jay, western tanager, and Oregon junco. In a nearby mature sequoia-mixed-conifer forest. Shellhammer (Hartesveldt et al. 1968) found the mountain chickadee and Oregon junco to be dominants, and the western tanager, jays, nuthatches, and flycatchers as subdominants. All the 12 common nonsequoian forest birds were present in these two sequoia areas, as were numerous other species.
Many birds tend to be more or less restricted to specific life zones, and hence the altitude at which a particular grove lies affects its avifauna to some extent.
Most of the 30 or more species listed in Table 3 are residents, breeding in and near sequoia groves. Over a period of time, one might count at least twice as many species if including the transients and occasional visitors.
The mammals of the sequoia forest communities are both common and numerous although, because many have nocturnal habits, park visitors see them much less than the birds.
The most common mammals are the deer mouse, gray squirrel, Douglas squirrel or chickaree, mule deer, golden-mantled ground squirrel, coyote, and black bear. They are found in virtually any Sierran forest.
The California ground squirrel, although normally much more abundant at lower elevations, is found as high as 6500 ft in the southern Sierra Nevada. It is relatively common in many of the groves, and appears to be more abundant in the groves lying within the national parks. Some claim that this results from the higher number of visitors and the correspondingly greater destruction of rattlesnakes which previously preyed more heavily upon the California ground squirrels in these areas. Others claim that the squirrels have always been numerous there, so the argument is far from settled.
TABLE 3. The more common birds within a typical giant sequoia grove classified by their feeding habits (Salt 1953, 1957; Kilgore 1968).
| Feeding habit | Species of birds |
| Air-soaring (raptors) | Goshawk, Red-tailed Hawk, Spotted Owl, and various other owls. |
| Air-perching | Western Wood Peewee, Olive-sided Flycatcher, and various Empidonax Flycatchers. |
| Foliage-insect | Winter Wren, Townsend's Solitaire, Golden-crowned Kinglet, Solitary Vireo, Warbling Vireo, Nashville Warbler, Western Tanager, and Black-headed Grosbeak. |
| Foliage-seed | Steller's Jay, Cassin's Finch or Purple Finch, and, in lower elevations, Band-tailed Pigeon. |
| Timber-search | Mountain Chickadee, Red-breasted Nuthatch, and Brown Creeper. |
| Timber-drill | Pileated Woodpecker, Yellow-bellied Sapsucker, Hairy Woodpecker, and White-headed Woodpecker. |
| Ground-insect | Red-shafted Flicker, Robin, Hermit Thrush, and Oregon Junco. |
| Ground-Seed | Mountain Quail and Rufous-sided Towhee. |
Most sequoia groves support populations of one or more species of chipmunks. The long-eared chipmunk occupies open, brushy, or rocky areas in the northern and central groves. In the same groves, the yellow pine chipmunk is found in open forest conditions. In the central and southern Sierra, these two species are replaced by the lodgepole chipmunk, normally found in groves in the Canadian Zone, and the Townsend chipmunk, usually restricted to those in the Transition Zone.
Occasionally, the golden-mantled ground squirrel shares a particular area with the chipmunks. Park visitors often confuse this squirrel with the chipmunk, but the former is considerably larger and lacks the stripes across the face which clearly identify the latter.
The one shrew which inhabits the dry forest floor of the Sierra's sequoia-mixed-conifer forest is the Trowbridge shrew, which is normally found in small numbers and, with its rather secretive activities, is rarely seen by visitors. This is true for most carnivores as well. While some visitors are fortunate enough to see an occasional coyote or even a bobcat, very few see the more elusive marten, the long-tailed weasel, or the mountain lion.
In the moist meadows and riparian communities throughout the Sierran mixed-conifer belt, one might find populations of mammals not normally found in the drier forested areas. The following mammals typify those habitats both within and outside of sequoian communities: vagrant shrew, water shrew, broad-handed mole, long-tailed meadow mouse, mountain meadow mouse, Botta pocket gopher, and raccoon.
Bats are more commonly seen during the evening hours, especially in the groves near the lower border of the Transition Zone. Although they are difficult to identify on the wing, the common species of bats in sequoia groves are as follows: little brown myotis, fringed myotis, California myotis, long-eared myotis, Yuma myotis, hoary bat, red bat, and big brown bat.
Only the hoary bat and red bat are tree-dwelling species, and may be found in the foliage of a grove in the daytime. The rest are cave-dwellers, or they roost in old buildings. All the bats are warm-weather visitors and migrate either into the Central Valley or southward with the onset of cold weather.
Of all the mammals in the sequoia forests, the chickaree or Douglas squirrel certainly plays the most significant role in the sequoia's life cycle.
The cold-blooded vertebrates constitute another varied group of animals not often observed by the average visitor. The term "cold-blooded," or its more recent synonym "ectotherm," refers to the amphibians' and reptiles' inability to maintain constant body temperature by themselves. Because they depend on mild climates and sunny areas, ectotherms are more numerous in the groves at lower elevations and the more open groves of the middle elevations.
The following reptiles and amphibians often inhabit the lower groves: Gilbert skink, western whiptail lizard, rubber boa, western terrestrial garter snake, and California mountain king snake.
Another group, more universal to most of the groves, regardless of elevation, includes: Eschscholtz's salamander, western toad, yellow-legged frog, western fence lizard, northern alligator lizard, and western rattlesnake.
One species, the sagebrush lizard, is found in the lodgepole pine-fir belt, chiefly in rocky, shrub-filled areas. Some sequoia groves, especially those protected by park status, shelter the rather rare western ringneck snake. However, most of these animals require either riparian or open, dry, rocky areas, and therefore are often missing from the larger groves. We might say that, in the large mature groves, the basic four are Eschscholtz's Salamander, the western fence lizard, the alligator lizard, and the western rattlesnake.
The food chain
Trees, shrubs, and herbs are the sequoia community's primary producers. These, plus the fungi, provide the nutritive base for the rest of the community. The green, photosynthetic plants support an amazing array of invertebrates, mostly insects, which are in turn consumed by a large variety of birds and some mammals. The reptiles and amphibians, although mostly insectivorous, play a relatively minor role in the food webs of sequoia groves because of their low numbers and the rather unfavorable environment for their existence.
Birds and a few mammals usually dominate the animal world of a giant sequoia grove. Most of the birds are insectivorous almost throughout the year although some of them switch to eating berries in the fall. The birds and the insect parasites are major agents in balancing insect populations. Where birds enter an area with large and growing insect populations, their territories tend to become smaller and their numbers increase. Which bird species abound in any one year, then, depends to a degree upon the species of insects in abundance. After a fire, for example, there is often a decrease in some of the ground-dwelling birds such as the mountain quail, Nashville warbler, and hermit thrush, resulting from the reduction of nest sites and of some litter-dwelling food organisms. In contrast, the populations of flycatchers and robins increase, in part because they are behaviorally adapted to more open forests (Kilgore 1971). Following the more intensive ground fires, after logging, or amid an outbreak of various forest insects, there is usually a decline among mountain chickadees nuthatches, creepers, woodpeckers, warblers, and tanagers. It is doubtful whether parasites and insectivorous birds together can contain insect populations at a sub-epidemic level, but they and other factors play a significant role in maintaining the balance.
A high percentage of the seeds of annual plants and many of the indigenous fungi serve as food for deer, and in particular for small rodents such as mice chipmunks, golden-mantled and California ground squirrels, so that their populations increase when these seeds and fungi are plentiful. The larger population reduces the next year's food supply, and the undernourished animals become less resistant to disease and starvation, and probably more vulnerable prey.
Relatively few vertebrates utilize the giant sequoia for food or shelter. A variety of foliage-feeders, such as the vireos, kinglets, and warblers, and air-feeders such as the flycatchers capture insects from or near the tree's foliage. Wherever the bark is thin, maturing sequoias often show the damage of sapsuckers, sometimes over much of the trunk in specimens whose bark is genetically thin. Stecker has found fresh sapsucker damage near the very top of each mature specimen he has studied. Heavy damage to younger trees sometimes occurs. Where the bark is literally peppered with sapsucker holes, the upper portion of the crown dies because of interrupted vascular continuity. Hartesveldt (1965) reports a 2.5-ft diameter specimen in the Suwanee Grove, Sequoia National Park, which has no fire scars, but whose trunk is covered with holes nearly to the ground and is a dead, dry snag in its upper 4 or 5 ft. Curiously enough, no record seems to exist that humans have witnessed a sapsucker in this activity on sequoias.
The thick, fibrous bark of older giant sequoias is often drilled by woodpeckers and is used for nesting, especially by white-headed woodpeckers and occasionally by flickers and a few of the perching birds such as nuthatches. There are reports of certain rodents, for example, chipmunks, using the dust accumulated in these bark cavities for dust baths. Some believe that the pulverized bark, rich in tannin, repels such common ectoparasites as lice and fleas. Other birds, such as flycatchers, will utilize these holes and limbs as nesting sites. The giant sequoia, however, is never exclusively sought out for food or nesting. The vertebrate that comes closest to reliance upon the giant sequoia is still the chickaree.
The role of the chickaree
The chickaree, (Fig. 34) is far from an exclusive devotee of the sequoia groves; it is in fact a resident of coniferous forests extending from the southern Sierra and San Francisco regions northward through the Cascades into Canada and Alaska. Its close cousin, the red squirrel, ranges from eastern Washington southward and eastward across the United States and Canada to the eastern seaboard.
The chickaree does have a peculiar relationship, however, to the giant sequoia. The general habits of the sequoian and nonsequoian chickarees are very much the same except for their specific feeding habits. Anywhere within their range, these hyperactive squirrels normally harvest the cones of pines and firs for the seeds' food value, removing the cone scales with their sharp teeth and dropping them wherever they happen to be feeding.
Outside of sequoia groves, they often build nests of twigs, grass, fungus, and bark for winter protection from the elements inasmuch as they do not hibernate. Thus, they must store food for winter use before the time of snowfall.
The study by Smith (1968) and the studies underway by Shellhammer and others reveal chickaree's year-round territoriality and an ability to maintain it by calling and chasing. This territoriality breaks down for mating purposes, but as soon as the year's brood can fend for itself, it is chased off by the parents to set up territories of its own. Variations in certain aspects of the chickarees' behavior in association with giant sequoias are not yet fully explained. For instance, they seldom if ever build external nests in sequoia groves, yet they are common especially in the more northern areas within their range. We know little about nest sites within sequoia groves, but there is evidence that the hollowed portions of tree trunks are sometimes used. Chickarees have been observed stripping the soft, fibrous bark of young sequoia trees, presumably to line nests. Chickarees conspicuously strip the bark of some trees, especially the lower portions of the trunks. Probably the most significant variation in the behavior of chickarees in sequoia communities is that they make the soft flesh of green sequoia cone scales a major food item.
Observers have long assumed that seeds were the main quest of the chickaree in cutting sequoia cones. Yet within their kitchen middens, remnants of individual seeds that have been bitten through indicate that their consumption is more likely incidental to their proximity to the more nutritious cone-flesh being eaten.
The number of cones utilized in a year's time varies greatly, depending on the supply of alternate foods such as the seeds of sugar pine, white fir, incense-cedar, etc., as well as the air temperature and the animals' sexual condition. Often a chickaree will harvest far more cones than it can possibly eat before the scales dry out and become unpalatable. When living in a given sequoia tree, a single chickaree may cut and eat as many as 3000-3500 cones per year. They are opportunists, however, and will switch to other, more productive tree species when the cones are at an edible stage, even though green sequoia cones are still plentiful. They also utilize fungi, hazelnuts, and a variety of other foodstuffs, but in a dense stand of sequoias, green sequoia cones appear to be their major source of food. Their diet varies proportionally to the diversity of the forest they inhabit.
Several years' observation in the Redwood Mountain Grove, where both chickarees and sequoias abound, indicate that the density of chickaree populations may vary greatly from one year to another. During years of high densities, the territories may be as small as 0.6 acre, and each animal seemingly requires at least one large sequoia tree and a tree of another species nearby, such as a large white fir or possibly a sugar pine. They seem to prefer the former as a nest tree, possibly because it is more prone to insect attacks and diseases, and therefore to drilling by woodpeckers, thus providing more nesting cavities than does the typical sequoia. In high density years, the hills and valleys resound with the squirrels' frequent high pitched calls and scolding chatter. In years of low density their home range may cover as much as 25 acres, with their defended territory near its center. During such years, the chickarees are relatively quiet, the only occasional tell-tale signs of their presence are the many freshly chewed cones beneath sequoia trees.
The chickarees' territorial defense merits further explanation. Defense is a type of agonistic behavior that contains elements of aggression (i.e., attack), retreat, or submission, Successfully evolved agonistic patterns seldom involve much actual contact or injury, if any, but rather consist of retreat, bluff, and easy surrender. Establishing a territory is an important phenomenon which among others provides its owner enough food upon which to live, but loses much of its survival value if the animals must kill one another to obtain and then protect their defended territories. Hence, the chickaree has evolved a behavior that relies on threats, reminders, and challenges in vocal form. The interloper usually does the same, and there is thus a relationship of emotional display rather than one of physical violence. In this manner, mutual recognition and respect allow them to live and let live, a requisite for species' survival.
The chickarees' territories change constantly in a given forested area. A chickaree may live in one sequoia for 3 or 4 years and prune a substantial portion of its total green cone crop. If they were longer-lived creatures, they might conceivably strip a tree of its younger cones, but such is not the case. Most chickarees mate at least once during their lifetime and drive away their young when reared, forcing them to take up territories of their own. When the parent dies, its tree may remain vacant for some years until, by chance, another squirrel claims it as home base. This results in a more-or-less continuous alternation or "rotation" of territories, insuring a more uniform usage of the tree's food supply over a long time (Fig. 42).
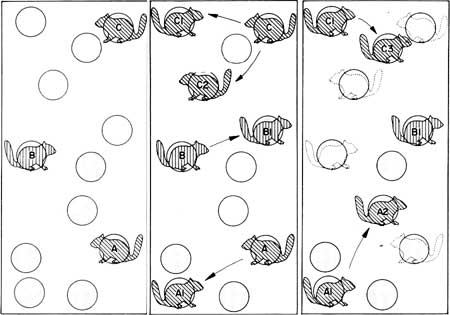
|
| Fig. 42. Hypothetical "rotation" of chickaree territorial use of sequoia trees for cone harvest. Flesh of sequoia cone scales, not the seeds, appears to be the energy source sought. Drawing by Ivan Linderman. |
The chickaree displays a decided preference for the younger green sequoia cones and generally eats only those that are between their 2nd and 5th years of growth. They seldom take the immature cones or those older than 5 or 6 years. The larvae of the beetle Phymatodes infest a high percentage of the older cones which are in varying stages of drying out on any given tree, and are perhaps too woody to be palatable to the chickaree. Cones stored by chickarees may remain green and edible for a year or more in the low, moist areas often used for storage. Those that are apparently cached in creeks are probably free-falls which have ended up in stream depressions quite by accident and most of which are unavailable for later use.
Some squirrels eat all their cones while in a tree's crown, and the seeds so released are spread over a greater area than those eaten on the ground (see "Reproduction"). Either way, seeds disseminated just before and during a snowfall may play an important role in sequoia regeneration. During most of the summer, the feeding of chickarees and beetles produces an almost constant fall of sequoia seeds, but many are lost to reproduction because of damage by insects, fungi, desiccation, and direct radiation.
Chickarees are small, very active mammals with very high rates of metabolism. Smith (1968) and others estimate that their basal metabolic rate is 1.76 times that expected from the weight-to-metabolism relation for most mammals. He suggests that, if the metabolic rate for most mammals were applied to chickarees, a male would still have enough energy to perform heavy labor 24 hours a day or to put on several grams of fat per day. A female could work just as hard and still supply energy to her young for their maintenance and growth. Chickarees do not hibernate and require considerable energy input every day of the year. In most parts of their range, they feed on large, high-energy conifer seeds, fungi, nuts, and berries. In the sequoia groves, however, the food is primarily the cone flesh, not the seeds.
Calorimetric or food value analyses of sequoia seeds and the outer cone material reveal that a chickaree would quickly starve to death eating sequoia seeds alone. However, they can and do eat enough cones each day to provide five or six times their basal metabolic rate. This figure was derived from cone counts around various trees multiplied by the average caloric value of a cone's eaten portion. The squirrels in our studies stripped approximately 3000-3500 cones per year, each cone yielding about 30 kg-cal. Thirty kg-cal equals the energy in two teaspoons of sugar, or a half slice of bread, or a medium raw tomato, While this diet may be greatly enriched by some sugar pine seeds in the fall, the cones are still the important overwintering food. Food that is four or five times their basal rate requirements appears more than adequate in the summer months, but not so in winter.
Red squirrels and probably chickarees have a rather high critical temperature of 68°F. The critical temperature is that below which the animal must expend energy beyond its basal rate to maintain a constant body temperature (Irving et al. 1955). This helps explain the correlation between temperatures and such evolutionary features in these two squirrels as nestbuilding, the caching of food underneath the insulating snow, daily foodgathering, etc., as well as the cutting of cones in large numbers and then making many trips to the ground to store them. But on this point, the chickarees of the Redwood Mountain and other groves begin to vary from red squirrels and chickarees from other areas. They expend great energy running up and down 300-ft tall sequoias. They apparently do not build large nests and, hence, probably expose themselves to the cold for the 20-25 minutes they take to eat each cone. This combination of food habits and behavior patterns in the long, cold winters may well push them to the limits of their adaptive abilities, and some starve or are weakened so that they fall prey to martens. Their life seems a biological compromise, with energy plentiful in the summer and often insufficient during the winter cold. Seldom, however, is a sequoia grove wiped clean of its chickarees—some of them are a bit stronger, healthier, quicker, more fortunately situated, or just luckier.
Some always survive to reproduce. In certain areas, they may have from four to five young and may breed a second time later in the summer, although apparently not in the southern sequoia groves, where they tend to have just one litter of from two to four young. Even so, they can build up rapidly in several mild years until the valley again rings with the territorial calls, and most sequoias are again having their cones cut. Not for long, however: an outbreak of plague, a long, cold winter, or heavier than normal predation, and down go their populations as the forest's complex ecological relationships bring them into bounds.
Invertebrate relationships
The invertebrate relationships of an ecosystem are very often relegated to a low priority of investigation unless there is clearly an economic reason to study them. Until recently, the giant sequoia was believed literally immune to insect attacks, and only 19 insect species were recorded utilizing the giant sequoia to complete one or more phases of their life cycle. Of the other invertebrates, virtually nothing is recorded except as they are related to commercially valuable tree species.
Although it may generally be assumed that invertebrates, especially insects, play significant roles in the interacting food webs of almost all ecosystems, some of the roles played remain unsuspected until such time as concerted studies are undertaken out of pure academic interest. Such was the story of the cerambycid beetle (Phymatodes nitidus), which probably plays such an important role in the reproductive life of the giant sequoia (see Seed Dispersal and Cone Fall).
The literature often states that insects have never been known to kill a mature giant sequoia. Although serious studies are lacking, this assumption may well be true. Actual observation suggests that insect depredations do not seriously affect sequoias after the second year of the seedling stage, and it is unlikely that insect attack alone causes their death. Yet many of these creatures contribute to the tree's loss of energy, and some to loss of structural support, all possibly aiding its eventual downfall. Damage or death of sequoias has been most noticeable not in native specimens in the Sierra, but where man has planted them in sites that were too moist. In such locations aegeriid or clear-winged moth damage has been significant in trees 5-15 years old. A colorful metallic wood-boring beetle (Trachykele opulenta) may kill specimens damaged by road cuts or the undercutting of stream banks. The larvae of these beetles, which remarkably can mature solely within the bark, usually girdle the tree by feeding within the nutritious inner bark area.
Insects are highly important in promoting the decay of dead trees and nonliving parts of all trees, including the giant sequoia: they make passageways enabling decay organisms such as fungi and bacteria to gain entry and thus reduce the plant remains to soil humus once again in the ageless cycle of decay and growth. The openings they make beneath the bark and in the wood tissues also permit entry of oxygen and moisture needed for decay.
The Phymatodes story is but one part of an intricate food web around the sequoia cone. Several species of parasitic ichneumonid, braconid, and chalcid wasps prey upon the Phymatodes larvae, thereby helping to check their populations. In fact, these parasites' specific searching behavior helps to identify the early work of Phymatodes larvae, which is impossible to recognize from the exterior, which, in turn, is an aid to the researcher in his attempt to find the early instar stages. A few insects use the old emergence holes and pupation chambers of Phymatodes for their brood-rearing, one of the more interesting being a snakefly (Aqulla assimilis) that lays its pale-green, banana-shaped eggs in the pupation chambers. Upon hatching, these highly efficient predaceous larvae will seek out invertebrate animal life for food, either within the cone or underneath adjacent loose bark.
During a cone's first year, when it is soft and green, it may be attacked—not always successfully—by a small gelechiid moth (Gelechia sp.). Only 0.50 inch across the wings, this moth deserves an award for its persistence. The larva, roughly one-sixteenth of an inch long, bores into the cone scale junctures, at which point it is usually repelled by the reddish cone exudate. The dead "pitched-out" larva is often visible within this small transparent mass of resin-like pigment. Less than 10% of these moth attacks are successful and most attempts abort during the first larval instar. We can recognize the moths' work by the browning of only one or two scales, although occasionally some mature larvae will cause up to two-thirds of a cone to turn brown. Internal inspection reveals mines three times the diameter of those made by Phymatodes, as well as distinctive fecal pellets and silk webbing.
In its larval stage, this moth also causes double cones by feeding in the cone's axial core and splitting it in half. Each half bows outward and distorts the shape of the maturing cone into an apparent double cone. Such dry cones can be broken easily into halves with the bare hands, unlike genetically formed "doubles."
The gelechiid moths' activity corresponds to how large a cone crop a given year produces. The new cone crop in 1970 was approximately four times that of the years immediately preceding. In fact, the upper third of the foliated portion of the Castro study tree contained nearly as many new cones as those of all other age classes combined, so that their total number on this tree rose from about 19,000 one year to more than 39,000 the following year. Gelechiid moth activities increased considerably at the same time. In the 1969 study, 1 of 11 cones showed evidence of gelechiid moth damage, while the proportion in the 1970 crop was 2 to 5, and in the upper 30 ft of the crown, 1 of every 2 was affected.
Brown, dried-out cones remaining upon the tree also harbor a small community of insects between the cone scales. Common in this community is a small lygaeid bug (Ischnorrhynchus resedae), about an eighth-inch long, that hides in the cone's openings when not feeding upon the juices of seeds and nearby foliage. It generally moves to the cone's center when an observer comes within about 4 ft of it. Psocids, or book lice, are also numerous within the diminutive labyrinthine passageways of the cone scales. Possessing chewing mouth parts, they cause minute damage to seeds that the opened cones may still retain, although investigations in the Castro Tree indicate that few seeds are really harmed, those still attached to the opened cones maintaining as much as 75% viability. This may result from strong vascular connections between the seeds and cone scales.
Several insect relatives that are predators also use the cones as a hiding place or as protective cover. The most interesting are the tiny pseudoscorpions, less than one-sixteenth inch long, which very likely feed upon the smaller book lice (psocids) also residing within the cone. Mites, jumping spiders (attids), and crab spiders (thomidids) are other arachnids which also live and feed upon the invertebrates associated with these cones.
An insect whose populations are apparently increasing in certain sequoia groves is the carpenter ant (Campanotus laevigatus). These black ants, often half an inch long, are the largest in the forest and live in colonies within the bark and dead wood of living sequoias, and in stumps, logs, and dead trees. Two or more colonies may inhabit one tree at a given time. They normally feed at night, but are sometimes seen fighting during daylight hours. More conspicuous than the ants themselves are the open-ended galleries which may honeycomb the tree's thick bark to a considerable extent (Fig. 43), and on the ground beneath is usually a mass of reddish sawdust chewed out by the insects' strong mandibles. The tunneling is expressly to make brood chambers for their young, the wood or bark removed in no way serving as food. Colony chambers may be as long as 20 ft, and the tunneled area a few inches to several feet wide.

|
| Fig. 43. Galleries of carpenter ants in bark of giant sequoia tree, Giant Forest. Recent interest in carpenter ant activities in sequoias suggests a possible relationship with increased human activities. Photo by Jack Hickey. |
Seasonal park naturalist Jack Hickey, of Sequoia and Kings Canyon National Parks, has long had an interest in the carpenter ants, and his preliminary study of this insect, particularly in Giant Forest, may show a possible link between the ant population increase and human activities. The carpenter ant scavenges on both plant and animal debris, including the food tidbits spilled by park visitors. The National Park Service has arranged for the University of California, Berkeley, to conduct in-depth research on the carpenter ant's relation to sequoia trees.
The damage is largely to the bark and of little direct consequence to the "host" sequoia. Yet the galleries do offer possible entry routes for other insects and decay organisms which may somewhat increase decomposition, and, perhaps more important, they provide access for air that would hasten the drying of bark and wood and thus increase the latter's inflammability. Dried wood becomes more brittle and the affected tree may weaken in time, fall, and die. Although the tunneling is generally confined to the base or near the bases of trees, a sequoia had its top knocked out by another falling tree in the summer of 1969, breaking off very near the site of carpenter ant tunnels about 90 ft above ground level.
Fire is apparently the nemesis of these insects. Even light ground fires seem readily to discourage ant populations by reducing their food supply. It has been suggested that their possible recent upturn is related to plant successional trends in the absence of fire, resulting in increased natural food. Present studies may eventually confirm this interpretation.
Other insects that create interesting food web patterns, while far less important in the sequoia's life cycle, are yet highly significant in the local faunal interplay. An insect not previously recorded as feeding upon the giant sequoia is a small green aphid (Masonaphis sp.) which is found throughout the larger trees, concentrated on the branches' tender tips in the lower third of the crown. It is also found on some of the seedlings along with another species of aphid. On a mature sequoia, these small sucking insects may number in excess of 100,000 individuals during late summer when they are at their peak. Each of the myriad tiny twigs on the tree may be host to ten or more per inch. Despite their juice-sucking activities, foliage discoloration is too slight to measure and photosynthetic reduction is believed to be almost insignificant. Nor do infested seedlings suffer from their feeding activities.
This rather large aphid biomass is preyed upon by the larvae of a syrphid fly (Syrphus sp.) that resemble elongate maggots. The visionless larvae find the aphids by probing randomly upon the branch tips where a female had oviposited her eggs among the aphids several days earlier. The larvae hang onto the narrow stems by means of posterior leg-like structures and thrust their bodies out into space after securing an aphid in their rasping mouths. Then the body fluids of the prey are sucked out. The effectiveness of the larvae in this blind search is borne out by research in which one individual was observed to have captured three aphids in one minute when the researcher removed the captive aphid by forceps each time one was caught. Normally, 3 minutes are required between captures
The predaceous syrphid flies also have their problems in this complex food web and many of them end up as food for the larvae of ichneumonid and braconid wasps which parasitize them. If they achieve adulthood, they are often eaten by a robberfly (Neoitamus affinis). These latter flies, which are highly predaceous, wait in ambush upon the ends of the sequoia's projecting branches. They capture adult syrphid or "hover flies" that are ovipositing along the outer margins of the foliage. They usually return to the site of the ambush to devour their fellow fly relative. The food web continues as the robber flies, in turn, are chased as potential food by several species of birds within the tree's crown.
In all, 151 species of insects and 37 arachnids are known to be associated with the giant sequoia in the sense that they use it to complete some part of their own life cycles. This organic web in living and on downed sequoias is highly complex. For some insects, the tree serves mainly as an unusual "high-rise" location for courtship: horse flies (Tabanus laticeps), bee flies (Villa), horn tails (Sirex areolatus), and even the tiny chalcid wasps hover over the tops of the tall sequoias as if to amuse or taunt an entomologist. In reality, they use this landmark to consolidate themselves for mate selection, a phenomenon called "hill-topping" in insects. Appendix IV contains the scientific names of invertebrates mentioned in the text. There are several species and subspecies of the snail Helm inthoglypta that are completely restricted to groves of giant sequoia trees in the southern Sierra Nevada (Rundel 1969). The relationship of the snail to the sequoias, however, does not appear to be established.
Although the invertebrate story for sequoia groves has been much increased within the last past few years, it remains fragmental. The sequoias that we studied intensively were all part of one large grove and were approximately at the same altitude. It seems reasonable to assume that studies at different altitudes and latitudes would further enlarge the scope and understanding of these relationships.
Finally, examination of the thick leaf litter layer beneath clusters of large sequoia trees reveals a rich invertebrate fauna co-acting with fungi, bacteria, and other decay organisms in the dynamic ecology of this layer and the soil. Studies in this area are almost totally lacking.
Competition within communities
In all biotic communities, each organism possesses its own distinguishing set of genetic traits. To be sure, there are similarities between one and another species but, like human fingerprints, no two are exactly alike. Thus, no two species of plants or animals react identically to a given set of environmental conditions, which insures that every species fills a specific niche in time and space, and the species' distinct genetic traits enable it to compete with other organisms present in that niche.
Green plants' success or failure in this competition depends also upon the availability of such essentials as solar energy, soil moisture, and nutrients. Competition within the plant world is strongly, but not exclusively, governed by the sunlight available at the photosynthetic or leaf surfaces. This is partly a function of stature, with the taller plants succeeding, and partly one of chlorophyll density within the leaves, as some shorter plants can compete successfully in reduced light. In both, the responses to light are inherited and determine vertical stratification of canopy levels as well as the sequences of plant assemblages over time. The new assemblages replace earlier communities as the plants' collective action and structure change the environmental conditions. Such sequential changes in plant communities are what we have called plant succession.
Plant succession
Plant successional trends are of two basic types: primary, which begins with the original breakdown of rock into soil materials to form a suitable substrate for the growth of higher plants; and secondary, or the sequential changes in plant communities following disturbances which destroy some of the plant cover, but which disturb the soil substrate less. For primary succession, it is sufficient to say that the soil base in sequoia groves is derived largely from granitoid and schistose rocks, and that it evolved over a very long period, antedating the giant sequoia's migration onto the western slope of the Sierra Nevada.
Secondary succession, however, recurs to some degree following such distrubances as fire, blow-downs, disease and insect epidemics, logging, etc. The degree of successional setback depends on the severity of the disturbance and the actual stage of succession at the time it occurs. The relatively short rotational sequence of secondary succession is of signal importance in succession theories and for vegetation management. Since disturbances such as fire appear to have occurred rather regularly in the natural sequence of events, the potentially long-enduring climax community was, in reality, somewhat rare in the mixed conifer forest of the Sierra Nevada before modern times. Fire was probably the most widespread and frequent disturbance; for the sequoia, it was perhaps the most beneficial factor leading to optimum conditions for reproduction.
With plant successional changes, there are recognized concurrent changes in both quantity and quality of the soil substrate. This is perhaps the more important in primary succession where sandy soils present a low cation exchange or nutritive capacity typical of soils low in the mineral clay fraction. Sierran granitoid soils, high in sand content, thus improve with age as the organic content increases. The humus performs a function similar to that of clay, enriching the developing soil, a factor of considerable importance in the seedling stage of many native plants. It plays a role in a plant's ability to compete with other plants in a community comprising several other species.
As soils increase in depth and organic content, this environmental change permits, through natural selection, other species of plants to invade, thus insuring also a change in the nature of both the litter layer and of the soil's humus content. The nature of the soil continues to change with each succeeding seral stage until at the climax stage of succession it comes into dynamic balance with the final breakdown of organic matter, and the soil depth becomes stabilized. Because of topographical variation, both plant successional and soil developmental sequences also vary greatly within short distances. This, coupled with set-backs by fire and erosion, forms both a mosaic of soil conditions and of plant community structure which thus gives the impression of a variety of climax plant associations. These are the often long-enduring sub-climax communities that typify the mixed coniferous forest region of the Sierra Nevada.
When intense fire sets back plant succession in the Sierra's mixed coniferous forest, the resulting plant community is usually dominated by a dense cover of shrubs. The species vary with the elevation and other geographical factors such as availability of soil moisture, steepness and aspect of the slope, depth of the soil, and, of course, the availability of a given species' seeds. Typical shrubs of this stage in the central and southern Sierra Nevada are deer brush, mountain whitethorn, small-leaved ceanothus, bitter cherry, manzanita, and wild goose-berry. These are usually mixed in a given locality, although one species may predominate, and may have an irregular sprinkling of herbaceous plants as well. There may also be a sparse and struggling overstory of almost any of the local tree species, perhaps relicts not consumed by fire or those seeded into optimum sites. Ceanothus not only seems well adapted to fire conditions, but it and lupine, which is a legume, have mutual relationships with nitrogen-fixing bacteria whose activities enrich the soil with nitrogen. Here, then, is a predetermined system of fertilizing the tree seedlings and saplings which follow in the succession. It is during this juvenile state that nitrogen is most necessary for their successful growth.
Where fires have been so intense as to destroy most of the tree vegetation, including the seeds, the shrubby stage may be prolonged for years because of the slowness with which trees invade from the edges and by which birds and mammals occasionally distribute seeds. Usually shrubs cover the fire-scarred land rather rapidly, soon providing more satisfactory protection against soil erosion until trees once again dominate the community.
The influx of trees may be of any local species, although the shade-intolerant ones tend to predominate. Their general characteristics cause them to dominate the early portion of succession. Because of their sensitivity to shade, these trees mostly possess the genetic trait of rapid vertical growth, enabling them to maintain their photosynthetic "factories" in as much sunlight as possible. Only thus do they survive. Such traits produce characteristics that man values—tallness and straightness. Trees that grow tall rapidly generally grow straight, reflecting the mathematical axiom about the shortest distance between two points. Tall trees require additional strength to maintain their very high crowns. Over the evolutionary years, natural selection has provided strength to the wood of most of these trees, which is reflected in the quality of lumber they produce.
In addition, these shade-intolerant species lose their lower foliage early in life as the leaves are shaded out, a process which not only continues as the tree gains stature, but is aided by the self-pruning of dead limbs. Lumber cut from these trees has fewer knots, a plus factor in the lumber trade.
In the mixed coniferous forest, the ponderosa pine, Jeffrey pine, Douglas fir in the more northerly groves, and the giant sequoia all qualify as shade intolerant, their genetic traits best insuring success during the period when sunlight can bathe their crowns. The assumption that sequoia, because of its size, is a climax stage species is clearly in error because it cannot reproduce itself in strong competition where shading may bring early death to many of its seedlings (see "Conditions for Seedling Survival").
Recalling that sequoia wood is relatively soft and light in weight, one may think it strange to include the giant sequoia in this early stage group. Of course, what it lacks in wood strength it makes up in sheer bulk, remaining indisputedly the tallest tree species in the Sierra Nevada. Its conspicuousness in climax communities composed predominantly of white fir does not identify it as a climax species, but merely as a relict which has continued to solve its light problems by keeping its crown higher than that of its associates. Indeed, if another species commonly grew taller than the sequoia, the latter would succumb because of its life-long shade intolerance. Thus, its genetically determined height has played a major role in the species' survival.
We pointed out that the giant sequoia was a prolific producer of seeds and that the conditions necessary for regeneration were rather exacting. Wherever fires, intense or not, burn through sequoia groves, they set back plant succession and to some degree help regeneration of sequoias and other early-stage trees. The sequoia's successful reproduction depends on distribution of undamaged seeds and success in seedling development. Because its competitive strength is roughly commensurate with that of the other early-stage trees, neither the sequoia nor the others have much opportunity to develop into pure stands, and rarely does one species predominate. Furthermore, where successional setback has been only partial, as it was so commonly because of past oft-repeated fires, other species that are somewhat more shade-tolerant also seed in, and provide additional competition for, the less tolerant species. The majestic sugar pine, largest of all pines, and the California incense-cedar seem to fall more properly into this category. Whereas sugar pine is generally spaced out, the incense-cedar often forms dense thickets which to some are not very aesthetic, restricting other more desirable species' reproduction, and furthermore clearly presenting a fire hazard. Like the sequoias, if they survive the hazards of their early years, they may grow into mature trees more pleasing to man. The old sugar pines are second in size and grandeur only to the giant sequoia and are the delight of visitors.
Where sequoia forest communities have escaped fire for a long period, the shade-intolerant species have multiplied greatly. The white fir appears to be best fitted for such an existence, and we find its seedling offspring surviving under the most shaded circumstances where other species are struggling or dead. Early photographs of sequoia groves indicate that white fir was considerably less abundant than it is now. Hartesveldt (1962), in an analysis of nearly 200 photographs taken in the Mariposa Grove between 1859 and 1932, noted an openness of the forest that is now scarce. In fact, the young white firs' preponderance today completely blocks many of the same views. These photographs further show that shrubs such as mountain whitethorn, manzanita, and bitter cherry were all more abundant in the past. Vankat (1970) has noted similar vegetational changes in the Sequoia National Park region that have resulted from the practices of early western man, especially grazing of domestic animals and practices of governmental land management agencies.
The changes just described were apparently caused by the advancement of plant succession attending man's highly successful campaign to prevent fires and to extinguish them as quickly as possible. This result, a stunning surprise to many, has created problems, some of which we have already discussed.
In the same context as the value of lumber of early stage trees, the climax species, white fir, is low on the scale. With no serious problem of light to solve, it does not attain the stature of others. Nor does its wood have the same degree of tensile strength; also, an unpleasant odor renders the wood less desirable as lumber. However, neither of these factors is directly related to its successional position. Tolerating shade, the lower foliage remains alive longer and, even when the leaves die, the dead limbs remain on the trunk, many of them remaining on old trees. This characteristic guarantees dry loose knots in the wood, again a liability for lumber use, and presents a somewhat cluttered view and obstacles to hiking. For the forest manager, it means increased fire hazard right to the ground level, and, in addition, the dense thickets of young white firs with their pitchy limbs and foliage can carry flames readily and rapidly at certain times of the year into the crown canopy.
While a serious problem for man, these features may be a well-ordered part of the natural diversity of life forms caused by successional stratification in time. We might regard them as ensuring that the earlier-stage plant species will again appear and not become extinct. While this may seem a bit contrary to older beliefs that fires are undesirable, it does make good sense if we can but screen out our own peculiar value judgments. Man has singled out and favored a few plants and animals to which he attaches monetary, practical, and aesthetic values. In attempting to increase their number, he has created circumstances that sometimes have actually reduced them. In fire prevention, man has inadvertently favored the less valuable tree species to the detriment of those with greater monetary value, so that the more valued early-stage trees have tended to disappear from the scene. Hypothetically, then, the sequoia could cease to exist sooner than if he (Western Man) had never entered the scene simply because of his well-intended protection of the sequoia.
It was stated earlier that man has had a tendency to see plant communities as assemblages of individuals at a given point in time. The comprehension of plant succession, although a step in the right direction, does not even necessarily imply an understanding of the community in its dynamic, sequential state of being. If one can visualize the entire spectrum of biota succession as an individual entity, as opposed to a static assemblage of plants, then perhaps there can be a closer appreciation of nature without the distracting value judgments of man. In nature, no one organism may be assigned greater or lesser value since each is only a functional part of the whole.
With dry limbs extending to or near the ground level, wildfire has an increased opportunity to destroy the dense canopy of the climax stage trees and permit the sun's rays to reach the ground. This, in turn, ensures that the shade-intolerant seedlings will have a better chance to survive to reproductive maturity. Again, because of the same dry limbs, even slow-moving ground fires can easily climb up the "ladder" into the crown of white firs which, though not necessarily destroyed by the flames, appear to be highly susceptible to desiccation and eventual death as a result of the heat. Thus, at least a partial opening of the crown canopy may result. While substantiating evidence is elusive, observation suggests strongly that these do occur. Following fires, there is generally an influx of those plants which were scarce in the later stages of the natural cycle of events, but were abundant when similar conditions prevailed earlier in the cycle.
Such knowledge becomes an advantage to man and aids greatly in the understanding of some of the problems inadvertently created out of ignorance of basic ecological principles. Man's changing attitudes toward the obvious implications of this knowledge, especially the fire relationships, will be discussed in the last pages of this book.
Where fires of lesser intensity burn through sequoia communities, the successional set-backs are also of a minor nature. The fewer trees of the upper crown canopy that are destroyed, the poorer the opportunity for shade-intolerant plants to reestablish themselves. With the disappearance of thick accumulations of leaf litter, small-seeded annuals increase greatly, but the shade-intolerant herbs such as the orchid (Goodyera oblongifolia) continue to maintain their populations at about the same levels as before. The giant sequoias will often germinate in great numbers after a light fire, but their chances for survival are much less under these more shaded conditions. Furthermore, the lightly burned soils remain more nearly unwettable than where incineration has been moderate to intense, and are therefore less satisfactory for seedling survival.
The undesirable changes due to the advanced state of plant succession as a result of fire control were clarified by Hartesveldt (1962) in a study on the effects of human impact upon the giant sequoias and their environment.
Human impact
At the outset of the reservation of our scenic treasures as national parks, very little concern was expressed over the deterioration that might occur from use by excessive numbers of people. The population of the United States was small in 1864 when President Lincoln signed into law the bill that set aside as a park the Yosemite Valley and the Mariposa Grove of giant sequoias. Because of the primitive means of transportation at the time, the demand for the use of such wild areas was but a tiny fraction of today's demand. The concept of ecology had not yet been formulated and the Congress and general public alike were simply and understandably not attuned to the concept of such problems, let alone to begin planning for them. The wording of the acts which established the various parks did mention preservation from impairment of the primary value being set aside, but it is wrong to assume that ecological connotations were the basis for the wording. Vandalism and thoughtless damage were doubtless the intent expressed in the acts. One person, however, had the imagination to project his thoughts far into the future and to foresee that the hundreds of visitors of his day would be the millions of a future day, and that mass use was likely to bring about changes of an undesirable nature to the park values.
His name was Fredrick Law Olmsted, creator of New York City's Central Park. He was selected as the first chairman of the board of commissioners of the new Yosemite Grant, operated as a state park. In 1865, having visited his new domain and interpreted the legislation that created the park, he issued a report that served as the basic guideline for management policy for the new park. In war-torn 1865, he showed rare understanding of the natural biotic communities, predicting the changes that we have now long recognized in popular park areas, Olmsted felt that the value of the parks would increase with growing population, and that from the very beginning, protective park management should serve posterity. Olmsted wrote (1952):
An injury to the scenery so slight that it may be unheeded by any visitor now will be of deplorable magnitude when its effects upon each visitor's enjoyment is multiplied by these millions. But again the slight harm which the few hundred visitors of this year might do, if no care were taken to prevent it, would not be slight if it should be repeated by these millions.
The full text of Olmsted's report is a literary masterpiece.
Not too surprisingly, in view of the then scarce ecological understanding, Olmsted's prophecy fell on deaf ears. His manuscript, too, was lost for many years.
In spite of some individuals' subsequent concern over detrimental changes, no overall policy advocated study or even careful observation to detect and rectify them. Apparently, it was only in 1933 that park biologist George Wright first included recommendations for such a policy in his book (Wright et al. 1933). Although not much came from Wright's ecologically oriented philosophy of park management, the thought persisted, and a publication of the U.S. Department of the Interior (1940) suggested ecological investigation as essential to the proper management of the biotic communities of national parks. Unfortunately, World War II delayed the implementation of such management programs for a good many years.
In an early, isolated instance, the National Park Service and other administrative agencies expressed concern over the effects of excessive tourist travel on the sequoias in the Giant Forest and in the Mariposa Grove, two of the most heavily used groves. This led to the employment of Dr. Emilio P. Meinecke, a Forest Service plant pathologist, who conducted a short survey in each of these areas to assess the damage done to sequoias and recommended ways to rectify the situation.
Meinecke's studies (1926, 1927) were disappointingly brief considering his findings and strong recommendations. The major point in each of his reports concerned physical damage to individual trees and the physiological damages he assumed to have occurred. There was no mention of community changes due to successional trends in the absence of fire, or of the relation between fire and sequoia regeneration. He was explicit, however, in his feeling that continued heavy use of sequoia groves was a threat to their future and that trees in the most severely compacted soils very likely would die. He did recommend restoring the parks to their original conditions, but without concrete suggestions for attaining the goal.
The two Meinecke reports, though not supported by extensive studies, were a beginning and led eventually to a 3-year study by Hartesveldt (1962) on the effects of human impact upon the sequoias in the Mariposa Grove. The results of this and subsequent studies in Sequoia and Kings Canyon National Parks shed considerable doubt on the old philosophy of noninterference by man to preserve specific biotic communities and their components, such as the giant sequoias. Moreover, they showed that Meinecke's fears were not well founded: human impact on sequoia communities was proven, but in forms not previously suggested.
Investigations revealed that soils were severely compacted in areas of heavy human foot traffic. Depending upon the weight of the body and the size of the foot, the latter's force in walking represents a pressure of 5-20 psi of ground surface. In areas where auto traffic had been permitted for years, the compaction was severe and the old roadways traveled by stagecoaches and wagons with narrow- and steel-rimmed wheels were still so densely packed after more than 35 years of abandonment that few woody plants had become established. But these narrow bands of compacted soil covered relatively small percentages of the larger sequoias' rooting areas.
In some regions with no pathways to channel visitors, the soils compacted by human traffic constituted as much as an acre of land. Around such popular trees as the Grizzly Giant, compaction greatly reduced water infiltration and the soil's capacity to absorb excessive surface flow. Such accumulated surface water created drainage channelways which then became gullies of erosion as volume and velocity of flow increased. In several places erosion had exposed the roots of sequoias to a depth of a foot and more, threatening the trees' future stability. Likewise, accumulations of water on impervious pavement had created similar channels to the side, some of them 3 and 4 ft deep. Yet to this date, no giant sequoia is known to have fallen as a result of this channeling.
Compacted soils were definitely shown to restrict penetration by roots, an effect that Meinecke believed very important. However, field studies of sequoia growth patterns revealed no sequoias tending toward death as a result, but rather an increased average annual growth rate of almost all sequoias growing in such soils. The soil profile of areas subject to extensive foot traffic showed a soil moisture regime almost universally more favorable than that in untrampled soils. Compaction to a degree restrictive to root penetration generally did not extend much below 6 inches in depth. The compacted layer seemed to form a protective cap which helped maintain the moisture level deeper down. Another important factor was the reduced competition for soil moisture by smaller plant species. The abrasion resulting from human trampling had largely eliminated these plants and consequently reduced most species' reproduction. Considering the relatively small volume of soil sufficiently compacted to be restrictive within a sequoia's rooting zone, very possibly the improved soil moisture at greater depths more than offsets the losses within the compacted zone. Whether affected trees adjust their feeder roots to greater depths, as has been suggested, is yet to be determined. Most significant is the fact that no record exists of a giant sequoia having succumbed due to compacted soil.
Changes in the chemical relationships of soils because of severe trampling fail to show any significance in statistical comparisons. The total organic content of the uppermost—or A1soil-horizon—has changed but little even in those areas where trampling has virtually eliminated the leaf litter layer. Reduction of basic soil nutrients was also light. The increased soil moisture at depths less than about 5 inches would undoubtedly have proven significant had a statistical comparison been made. Although these changes were of no consequence to the larger sequoia trees, they did have a profound effect upon the smaller herbaceous plants and seedlings of both trees and shrubs. Surface soils were powdery dry by late summer and, in these circumstances, sequoias do not seem to produce seedlings at all.
Two other great concerns were the effects of road-building activities, namely, road ballast and pavement placed over sequoia roots, and the cutting of roots from the side of large specimens to accommodate road cuts. Deep cuts made close to a trunk, on occasion, eliminated as much as 35% of the entire root system.
Pavement which covers rather large portions of a tree's entire root system produces growth increases exceeding those mentioned earlier in compacted soils. Not only does impervious pavement eliminate plant competition, but surface evaporative losses are also reduced to a minimum leaving the "relict" sequoia a greater supply of soil moisture that enables growth to continue beyond the normal growing season. In effect, then, the growing season for such trees extends late into the fall and, in some cases, until freezing weather commences in December. One such specimen, the Sentinel Tree, situated between the Generals Highway and the Giant Forest Village parking lot, has about 75% of its root system covered with pavement, causing great concern until increment borings showed the tree growing nearly 50% faster than before the addition of pavement. Vigor of growth, however, may not be the final criterion of man's influence on such trees. The effects have been measured over a small percentage of the tree's total life, and these changes may eventually prove harmful.
Where roadway construction has cut away roots, annual increment to the trunk has declined significantly immediately after severance because of reduced delivery of soil moisture and nutrients to the crown's photosynthetic factory. Root removal would seem to reduce proportionately the crown foliage, just as large burn scars do, yet the trees so affected still have healthy crowns. And, curiously enough, the growth patterns developed during the years since the roots' removal show a gradually increasing growth rate even though the total rooting area has not seemed to increase. This may be due to proliferating feeder roots in the reduced root zone, or to greater lateral translocation of photosynthate in the trunk. Here again, no sequoia has apparently fallen or otherwise died because of either of these disturbances by man. However, considering the regulations governing the management of sequoias, this should not be regarded as license to treat them in such a manner.
The most profound change since the advent of western civilization had hitherto not been suspected, although Joha Muir's observations on the relationships of fire could well have permitted him to recognize it. The change resulted from man's benevolent regard for the great sequoias and his abhorrence of wildfires. His successful program of fire prevention and suppression was more than admirable; yet he had failed to recognize the disadvantage he had prescribed for the sequoia and other early-stage plants whose reproductive success was diminishing in competition with shade-tolerant species. The virtual elimination of sequoia reproduction remains not too serious a problem for species survival. A tree whose life span is 2000-3000 years or more can maintain the species by reproducing under favorable conditions after several centuries of failure. But the circumstances man has created have possibly a more profound effect than he knows, and consequently the studies we have described were undertaken to better understand the relationships of fire to the giant sequoia. Despite considerable knowledge at this time, we can undoubtedly learn much more. Our final section will discuss further these relationships.
The current assessment of changes to sequoia ecosystems by man is considerably different than it was earlier. The direct changes are sometimes expressed as damage to the ecosystem; yet these damages are restricted to relatively small areas and do not seem to affect adversely the larger specimens. The more subtle, indirect effects pose more difficult problems. However, management procedures can control most of those regarded as detrimental to man's interests, and many necessary changes now being implemented will better ensure the future integrity of the species.
| <<< Previous | <<< Contents >>> | Next >>> |
chap6.htm
Last Updated: 06-Mar-2007