|
VI. Types of Forest History and Dynamics
The lodgepole forests in Crater Lake National Park have several
apparent types of stand history: Type 1) Those seral lodgepole forests
which are rapidly replaced by fir and hemlock need to be considered as a
part of the larger complex of fir-hemlock forests. At any one time, part
of this complex is in mature fir-hemlock forest, part in seral lodgepole
stands, and part in transition (Fig. 1).
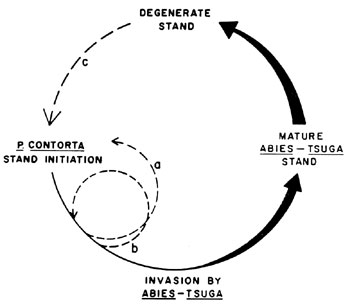
Figure 1. A proposed model of stand development in areas supporting
seral P. contorta forest on pumice soils. Heavy arcs indicate
phases in which intense fires are unlikely. Solid fine curves indicate
phases in which intense fires are more probable. Broken lines signify
fires. Fire types a and c are intense enough to initiate a new P.
contorta stand. Fire type b may initiate a new age class or only
burn the understory and tree reproduction. (from Zeigler, 1978).
Intense fires, which kill the fir and hemlock, create new lodgepole
forests, which may then develop into fir-hemlock with time. The
lodgepole reproduces poorly in these dense seral forests and beetle kill
hastens its demise. Litter is heavy; a reburn would probably kill most
of the lodgepole but would also produce a new lodgepole forest. In most
stands of this type, the major age class originated after white man
arrived. From historical records it seems that a larger proportion of
the area which can potentially support fir-hemlock is now in lodgepole
than there was in primeval conditions. This resulted from the many fires
in 1855 to 1900, some of which burned in mature forest. Type 2) Some
areas which can support fir-hemlock are invaded by trees only slowly
after forest destruction by fire, probably due to a relatively dense
herbaceous cover. Gradually the lodgepole pine increases in number, with
most reproduction being near older trees, forming islands of forest in
which fir and hemlock become established. These tree islands gradually
spread into the meadow between, and, probably after two or more
generations of lodgepole without a major disturbance, a closed forest
may form. In the meantime, however, some individual tree islands were
probably destroyed or thinned by local fires and by bark beetles,
delaying forest closure. Tree growth is very rapid once trees finally
become established, and they reach beetle-susceptible size at a
relatively young age. Type 3) In some lower elevation areas, contiguous
to the ponderosa pine forest, periodic ground fires probably maintained
a mixed forest of lodgepole, white pine, fir and hemlock. Fuel loads are
low, and the burns were probably small or patchy, and of low intensity.
Large trees, even lodgepoles, were scarred without dying, but most of
the reproduction in the burns would have been killed, and lodgepole
reproduction increased in the openings. One fire, which increased the
fuel load as dead trees fell, probably led to a greater chance of a
later reburn there. If a long enough time passed without fire, an
intense fire, killing most trees, could probably have been supported by
the accumulated fuel. Lodgepole re-invasion could have led to another
forest maintained by periodic ground fires. The burning interval between
the only two fires recorded on scars was 30 years, but now, 80 years
later, fuel loads still appear too low to allow other than patchy fires.
Type 4) Some areas appear to be lodgepole pine climax, where fir and
hemlock rarely establish. In the better sites, an open or patchy
lodgepole forest gradually may become a quite dense thicket, stopping
lodgepole reproduction at least in spots. In the patchy phase fires were
probably small or of low intensity due to discontinuous or light cover
of litter. These small fires and beetle kills delayed the development of
closed forest. After the forest closed, and most likely following heavy
beetle kill, intense fires occurred, killing all trees and beginning the
cycle again. After 75 years of fire protection, some of these forests
have developed densities and fuel loads which are very conducive to
intense fire and probably equal or exceed the maximum present under
primeval conditions. Type 5) Some climax lodgepole forests are very
sparse in all layers, and grow in habitats which appear incapable of
supporting denser forest. Fire would be confined to very small patches
of continuous fuel. It seems unlikely that extensive fires of any type
could have been supported. Reproduction occurs more or less
continuously. Beetle kill or local fires remove the older trees;
however, some with heavy mistletoe survive longer than any trees in
other communities. In these forests, fire effects appear minor, the
stands are in more or less a steady state, and further stand development
will be a process of primary succession, occurring only over many
generations of trees.
VII. Plant Communities in Lodgepole Pine Forest
Eleven communities were defined in the lodgepole pine forest. These
communities are named after the apparent climax tree species and
dominant shrubs and herbaceous species. A key for the identification of
the communities in the field accompanies this report as Appendix B. The
general distribution of these communities is shown in a type map
(Appendix C). We strongly urge that the map be used only for general
orientation and that the key be used when deciding management policies
for any particular location in the field.
In general, we found that no one community can be said to result
entirely from man's activities, though some types apparently prospered
as a result of the numerous fires that accompanied the white ' s arrival
in the area. One community appears to have experienced fairly frequent
ground fires, as well as quite severe fires. Contrary to the popular
belief that lodgepole pine is usually seral, we have found three
communities where lodgepole pine is the only tree even in old stands,
and is reproducing in large numbers.
Brief community descriptions are given below. Accompanying data are
presented in Appendices E, F and G. Included in these descriptions are
what we believe to be the disturbance histories and the consequences of
a fire at the present time. For more complete community description and
the facts upon which this summary is based the reader is referred to
Robert Zeigler's Masters Thesis. The number in parentheses beside the
community name corresponds to the number of the community on the type
map (Appendix C).
(1) Incense Cedar/Manzanita
This community is found on steep rocky slopes along Annie Creek
Valley. The vegetation includes sparse forest with numerous herbs and
shrubs growing among the rocks. Ages indicate that the fires that
probably infrequently burned this type likely originated in lodgepole
stands downslope from it.
(2) Lodgepole Pine/Bitterbrush/Sedge
Stands of this type are found in the northeast quarter of the Park
between Sharp Peak and Desert Creek at elevations between 1650 m and
1750 m. The herbaceous vegetation is similar to community 3, with the
addition of a shrub layer of bitterbrush and, to a lesser extent,
rabbitbrush goldenweed. These generally open stands are composed of
almost pure lodgepole pine. The apparent successful reproduction by
lodgepole pine in the absence of fire, and that all charcoal is from
lodgepole pine, indicate that this community is a true lodgepole
climax.
There is evidence of past mountain pine beetle activity, though
litter accumulation is still fairly light. Because of the patchy nature
of the ground cover, light ground fires were probably not extensive.
Fairly infrequent intense fires probably recycled the stand after heavy
fuel buildup. Most of the areas occupied by this community are probably
incapable of supporting either kind of fire at present.
(3) Lodgepole Pine/Sedge-Needlegrass
This community is found on flat areas and depressions with deep
pumice and/or scoria deposits at elevations from 1570 m to 2000 m. The
largest examples are in Pumice Flat, around the Pumice Desert and on the
west side of Sand Creek. The ground vegetation in this type is
characteristically depauperate, consisting mainly of a sparse, patchy
cover of sedges and grasses. There are very few, if any, shrubs. Though
there may be some hemlock and white pine near other communities,
lodgepole pine is usually the only tree species present in all layers.
Therefore, this community is considered a true lodgepole climax.
Most stands were extensively thinned by mountain pine beetle
epidemics in the first half of the century. The thinned stands support
relatively vigorous lodgepole regeneration. Most older trees are
severely infected with dwarf mistletoe. Considering the present fairly
abundant reproduction, this will probably lead to stands being heavily
infected with dwarf mistletoe; this was likely also the case in the
primeval forest.
Stands in this community probably burned only rarely and then only
over small areas The litter accumulation, even after 70+ years without
fire, is very patchy with islands of heavy fuels separated by large
areas of mineral soil Openings in the stand permitting lodgepole
regeneration probably resulted from beetle kills. Any fire starting in
this type would probably be quite small--limited to one snag or a
locally heavy collection of litter. That fire was relatively unimportant
in the community in pre-white man times is further supported by the
great ages of the stands and the scarcity of charcoal on the forest
floor. All charcoal is from lodgepole pine.
(4) Lodgepole Pine/Sedge-Lupine
This third lodgepole pine climax community is found in extensive
areas about the Park. It is most accessible on the west side of Sand
Creek. Other large stands may be seen northeast of Cascade Spring,
southwest of Sharp Peak, west of Timber Crater and southeast of Bald
Crater. Stands of this type, found between 1700 m and 1980 m, are
recognized by the presence of pine (Anderson's) lupine. In some areas
goldenweed and squaw current may be present.
Areas supporting this type were probably visited by intense fires in
the past as suggested by the presence of only one or two age classes in
all but 2 of 13 sample plots. Recent high bark beetle activity and
apparent ice breakage have led to very heavy litter accumulations in
some areas. This natural buildup has been increased by locally dense
reproduction, resulting in areas of apparently very high flamability.
These areas are also characteristically severely infected with dwarf
mistletoe. It seems likely that areas such as these would have burned
before now without fire suppression. Fire in the area would probably
result in nearly 100% tree mortality with a short term reduction in
fuel. Dwarf mistletoe in the stand would be eliminated or greatly
reduced. As mortality from the fire fell the fuel load would again
increase. Another fire, consuming this post-fire fuel and corresponding
reproduction, would probably permit the establishment of a stand of
vigorously growing trees in an open meadow-like environment.
The closed, highly flammable areas of this community are found
between the North Entrance Road and Timber Crater and at the
southeastern end of the Pinnacles Valley. Open stands, whose origins are
likely those hypothesized above, are found in the upper western
Pinnacles Valley, the area southwest of Sharp Peak, and west and north
of Desert Cone.
(5) White Fir /California Brome-Lupine
This community is found only in a small area northeast of the
Panhandle and west of Sun Creek at elevation 1460 m. White fir is the
dominant tree in the understory. There is extreme accumulation of litter
from past bark beetle epidemics in some areas. Age data indicate that
this type existed prior to the white man's arrival in the area.
Following 1855, fires may have increased the area occupied by this type.
A fire at present would probably destroy most of the stand in some
areas, with lodgepole pine re-establishing itself following fire.
(6) Subalpine Fir/Collomia-Peavine
This community is found in very wet areas near the headwaters of
Bybee Creek and Copeland Creek at about 1700 m. The community is best
distinguished by the presence of collomia and peavine, though very wet
sites may contain a rich flora. Lodgepole pine grows very rapidly on
these sites and both Shasta red fir and subalpine fir occur. The
dynamics of this type are probably quite similar to the subalpine
fir/goldenweed/aster-blue wildrye type (no. 7 below), though tree
invasion is even slower because of intense competition with herbaceous
species.
(7) Subalpine Fir/Goldenweed/Aster-Blue Wildrye
This relatively lush seral community is found between 1540 and 1920 m
in the vicinity of streams and at the base of steep ridges. The most
extensive stands are on the west slope of Mount Mazama, Munson Valley,
and along upper Sand Creek. Smaller stands occur near Sphagnum Bog,
Crater Springs and Pole Bridge Creek. Floristically, this type differs
from others in the presence of Cascade aster, blue wildrye, Green's
rabbitbrush and/or Rydberg's penstemon. Subalpine fir is also present in
almost all areas. Rather than being a true forest, the community is a
forest-meadow mosaic. Patches of relatively dense trees of all sizes are
separated by relatively lush meadows of lupines, grasses and sedges. The
islands of tree reproduction appear to be slowly spreading into the
meadow areas. Heavy litter accumulations occur only in the tree islands.
In older, nearly closed stands, such as those found in upper Munson
Valley, tree mortality from mountain pine beetle has been and continues
to be quite high among older, larger trees.
Most of these areas were burned before 1900 by ranchers, to improve
grazing for their herds. Age analysis indicates that most of the west
slope stands are of post-white man origin while those in Munson Valley
contain pre-white man age classes. Charcoal data indicate that some
earlier stands contained predominantly fir and hemlock. Fires in this
type, at present, would probably be limited to a few tree " islands'"
and the intervening meadow-like areas. In the primeval forest, intense
fires through nearly closed forests of this type probably resulted in
very open forest-meadow mosaics. These mosaics gradually closed over
several generations of trees, with closure retarded or temporarily
reversed by periodic light or small fires. The closed forests either
burned again or developed to pure fir-hemlock stands.
(8) Shasta Fir-Mountain Hemlock/Sedge-Lupine
This widespread seral community is found between 1690 m and 2080 m
through out the Park. Extensive stands may be found in the northwest
quarter of the Park, on the slopes of Timber Crater and in Castle Creek
Valley. This community is recognized by the presence of pine and/or
broadleaf lupine in an understory of conspicuous and apparently vigorous
fir and hemlock reproduction.
Bark beetle activity and breakage at galls on the main stems of
trees have contributed to a heavy accumulation of lodgepole pine litter.
Fires in this community would probably result in nearly 100% tree
mortality and a post-fire forest of lodgepole pine. However, litter
loads would again be high within a decade or two after the fire as
fire-killed trees fell.
Age analysis of stands comprising this community reveals that only
half of the stands contain trees which germinated before 1855. Charcoal
from some of the stands indicates that the sites were occupied earlier
by fir and hemlock forests. In addition, many stands contain old,
unburned logs and stumps that were obviously quite old firs and hemlocks
from a previous forest. Some stands contain surviving large trees of
these species. Other stands of almost pure medium-sized fir and hemlock
contain a few very large lodgepole pines and have considerable lodgepole
mortality on the forest floor.
These data and observations in this community suggest that:
1) A natural cycle exists where lodgepole pine forests are created
from mature fir-hemlock forests by fire. Lodgepole pine forests created
in this manner may be maintained as lodgepole by repeated fire for a
period of time before developing to fir-hemlock again (Fig. 1).
2) Fires caused by white man in the late 19th century increased the
area of this community and created areas of lodgepole that were
previously in fir-hemlock. Thus, the area of this community is larger
now than in the primeval situation.
(9) Mixed Conifer/Manzanita-Bitterbrush/Sedge
This community is found only in steep slopes northeast of Mazama Rock
at elevations around 1770 m. It is similar in structure and composition
to the Mixed Conifer/Manzanita community. It apparently experiences
periodic ground fire. Severe fires are probably infrequent.
(10) Mountain Hemlock/Grouse Huckleberry
This seral community, found between 1600 m and 1770 m, is recognized
by patches of grouse huckleberry in an otherwise depauperate understory.
Tree reproduction is mixed hemlock and fir with the former usually
dominant. The litter accumulation, age structure and apparent history of
this type are similar to the Fir-Hemlock/Sedge-Lupine community (number
8).
(11) Mixed Conifer/Manzanita
This is one of the communities of lodgepole pine that probably
experiences fairly frequent ground fires. It grows in small areas
throughout the Park between elevations of 1570 and 1900 m. The sparse
understory is dominated by pinemat manzanita and/or greenleaf manzanita.
Tree reproduction is well represented by Shasta red fir, western white
pine and lodgepole pine. Ponderosa pine may be found in stands on the
east side of the Park. A sizeable stand is found along Highway 62 north
of the Panhandle. Other stands may be found along the East Fork of Annie
Creek, the east side of Sand Creek, northeast of Mazama Rock and west of
Bald Crater.
These stands are typically quite old and heavily infected with dwarf
mistletoe. Bark beetle mortality is apparently continuous. Many trees
exhibit fire scars with the interval between scars on white pine being
between 30 and 40 years. This community probably experiences several
light fires be tween the infrequent severe fires which would be
responsible for stand destruction. These light, patchy fires would allow
continued reproduction by lodgepole pine.
|
