|
NATIONAL PARK SERVICE
Giant Sequoia Ecology: Fire and Reproduction |

|
CHAPTER 7:
THE ROLE OF INSECTS IN GIANT SEQUOIA REPRODUCTION
Ronald E. Stecker
Introduction
The sole reference to insects affecting giant sequoia reproduction prior to this study was a brief notation by Fry and White (1930) stating that cutworms damaged plantation stock. The present study, however, revealed as early as 1966 that several arthropods, especially insects, have an impact upon the reproduction of the giant sequoia. The discovery in 1968 of the cone activity of a small cerambycid beetle, Phymatodes nitidus, led to studies showing the importance of this insect in the release of seeds from giant sequoia cones (Stecker 1973).
The objectives of this part of the investigation were twofold. The first objective was to measure insect-related losses of seedling sequoias and the second was to study insect activity in ovulate cones in the crown of living trees. Data were collected in respect to the latter objective to determine the extent of damage to cones, the age classes of cones used by the cone beetle, and the effect of insects upon the dispersal of seeds and reproductive potential of the giant sequoia.
The four study areas described in Chapter 2 were used in this study, as well as special plots in or near the Redwood Mountain Grove which are described in the following section. Two mature giant sequoias were rigged with an electric elevator for in-crown investigations, the first such research to be conducted 200 to 300 feet above the ground in this tree (see Chapter 6).
Methods
A census was made of hypocotyl loss, defoliation, and damage from sucking insects of seedlings in the study plots and along several arbitrarily selected transects. The major emphasis of this study was given to measurement of cone insect activity in the crowns of pole size to mature 93 m (300 ft) tall sequoias.
Artificially seeded plots
Seedling beds were artificially established in Redwood Canyon and on Redwood Mountain to assess insect loss of young sequoias from germination through the first seasons growth. Three plots of 3m x 3m (10 x 10 ft) were established in the fall of 1967 and early in the summer of 1968 in Redwood Canyon at sites of different successional stages. Ten plots were established on Redwood Mountain on 1969. Finally during the summer of 1970, 28 random, uneven-sized groups of natural seedlings were marked in the latter area. The seedlings in these plots were studied primarily for insect activity and not survival as such.
Plots for 1968. and 1969 were established upon diverse sites. In each case litter and duff were removed to expose bare mineral soil. Sixty grams of seeds (approximately 12,000 seeds) were then lightly raked into the top 3mm (1/8 in) of soil and tamped in by foot and the end of a rake. Plot 1 was within the southeast boundary line of Trail Area (Fig. 4) adjacent to the road where no natural seeding had taken place. Trail Area had been manipulated by fire in 1965. Seeds were sown during the late fall of 1967 to minimize loss of seed viability due to desiccation. Plots 2 and 3 were located in Redwood Canyon, 0.4 kilometer (1/4 mile) south of South Area. Duff was removed to expose mineral soil and the plots were provided with approximately 1.3 cm (1/2 in) of water per week during the growing season by means of perforated five gallon tins and by fire-control pumper truck. Plot 2 was a shaded depression surrounded by white fir.
Seeds were planted the last week of November 1969, just before the winter snowfall, in 10 prepared seed beds measuring 3 m x 3 m (10 x 10 ft). These plots were numbered from 4 to 13. Seven of them were situated within the 305 m (1000 ft) wide 1969 National Park Service environmental burn along the ridge of Redwood Mountain, which extended 1.6 kilometers (one mile) to the south from Redwood Saddle (Fig. 4). Three control plots, cleared to mineral soil, were established outside that burn.
Naturally seeded plots
It was noted in 1970 that seedling establishment from natural seedfall of the 1969 environmental burn atop Redwood Mountain was at least four times greater than that found in the prepared seedbeds in the same area. This was surprising considering the hand seeded rate of approximately 12,000 seeds per 3 m2 (10 ft2) plot. The apparent heavy natural seedling establishment was in part a result of cones that had been killed by rising masses of hot air during the environmental (fuel reduction) burn (Kilgore 1970). Seed drop was apparently from cones from the 30 to 70 m (100 to 225 ft) level of these trees. Study efforts were then concentrated on this area of naturally seeded reproduction, where seedling density reached a maximum of 82 individuals 1 dm2. Twenty-eight plots, each with 10 tagged randomly selected seedlings, were picked from within the 305 m (1000 ft) wide area. These plots were numbered from 14 to 41.
One meter wide transects were also established at Trail and South Areas to assess the loss to seedling insects during the summers of 1968 and 1969. The transects were laid out on diagonals with sampling taken one meter in every 10 meters.
In-crown sampling methods
Techniques described in Chapter 6 were used to gain access to the cone distribution in a number of study trees (the Albina, Castro and in other free-climbed trees). Approximately 4,000 cones were removed from these trees, opened and cut in quarters. Most cones were sliced using a large Corona pruner with a 10.2 cm (4 in) blade with one of the 74 cm (30 in) handles attached by clamps vertically from a heavy, broadbased platform. The free handle provided sufficient leverage for clean cuts of both green and brown cones. Heavy knives and meat cleavers were excluded as too dangerous to use repeatedly, and because they lacked the precision of the oversize pruner. Cones were bisected lengthwise twice, providing quarters of cones for easy examination. Most quarters were observed under a binocular microscope using 15 power magnification.
A major emphasis in this study was focused on the effects of insects upon the ovulate cones of the giant sequoia. The persistence of cones in this species allowed for precise and quantitative observations on insect activity relative to location, age and spatial distribution of the cones. This necessitated a total census of the cone load in at least one tree. A mature sequoia usually contains from 10,000 to 30,000 or more cones. They are unevenly distributed in a vertical and horizontal manner. The distribution of cones in the Castro Tree was carefully established by hand counting in order to avoid the high error associated with visual methods (50% or more). The cone count and mapping was made of the entire Castro Tree in August 1970. Age classes of cones were tagged using different colors of plastic "flagging" tape for each respective year.
New cones which are pollinated during early spring are approximately one-fourth smaller than the second year cones. These "yearling cones" are light lime green and are the ones most easily observed from the ground because of their apical position. Older age class cones are usually obscured by foliage. The first year cones are also the only ones with a 2 to 4 mm aristate tip (bristle) distinctly projecting from the center of each scale's umbo (Fig. 48). This tip dries, shrivels and starts to disappear by the second year. Thus, first and second year cone classes were easily identified for census purposes.

|
| Fig. 48. First year cone which shows aristate tips of scales. |
Selected cones with known insect activity were encased in live traps of two types; one, a nylon mesh cylinder with cord ties at each end, the other, a 7.6 x 12.7 cm (3 x 5 in) cotton mailing bag with a drawstring end. Cones selected for rearing insects were housed in number eight Kraft paper bags. The bags were used even though a few beetles bored out through the paper. Cardboard boxes and large metal cans were tried, but proved unsuccessful because of the development of mold on the moist cones.
Results
Insects appear to play a major role in the reproduction of giant sequoias by releasing seeds from cones in standing trees and causing loss to seedlings.
Seedling loss to insects
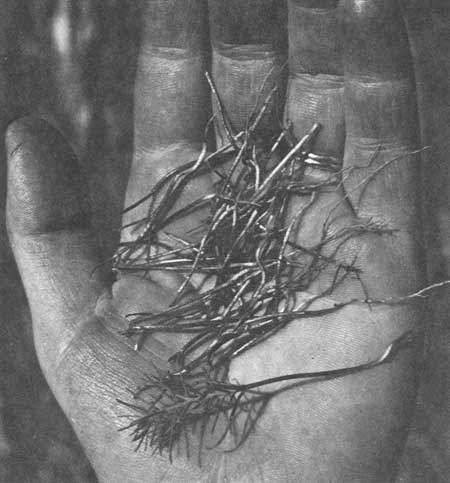
By August of 1968 6% (52 of 871) of the cotyledon stage seedlings in plot 1 had had their tops chewed off. Plot 2 (where 206 seedlings developed) had 3.4% (or 7) of the seedlings lost to chewing insects, while plot 3 sustained a 4.2% loss or 41 out of 966 seedlings to chewing insects. Deer ate 26 seedlings one night in what appeared to be four random browsing attempts in a line across plot 3. The transects in Trail and South Areas revealed 3.7% and 4.3% mortality respectively from insect activity. Fig. 49 shows severed hypocotyl loss to insect feeding, which can be easily distinguished from deer browse by the mandibulate markings.
|
| Fig. 49. Severed hypocotyls of giant sequoia seedlings due to insect feeding. |
Insects responsible for the above chewing loss were early instars of Pristocauthophilus pacificus, a camel cricket (Gryllacrididae), and the larva of the geometrids Sabulodes caberata and Pero bebresarius. The former, an orthopteran, is a nocturnal feeder in contrast to the diurnal feeding of the caterpillars.
Inspection of the prepared, hand-seeded plots numbered 4 to 13, in the early summer of 1970, revealed that 17.5% of the seedlings were completely chewed off (hypocotyl severed) (Table 12). Another 34.2% showed chewing damage to either cotyledons or needles. Control plots did not contain any seedling development.
Table 12. Insect damage to giant sequoia seedlings in prepared seedbed, Redwood Mountain—1969.
| Plot numbera |
Number of seedlings |
Hypocotyl severed |
Cotyledons chewed |
Leaves chewed |
Total % damage |
| 4 | 22 | 6 | 3 | 5 | 63.6 |
| 5 | 12 | 0 | 1 | 4 | 41.7 |
| 6 | 47 | 8 | 14 | 6 | 59.6 |
| 8 | 15 | 3 | 1 | 3 | 46.7 |
| 9 | 8 | 0 | 0 | 2 | 25.0 |
| 11 | 28 | 8 | 2 | 6 | 57.1 |
| 12 | 17 | 1 | 2 | 2 | 29.4 |
| Total | 149 | 26 | 23 | 28 | |
| % insect activity | 51.6 | 17.5 | 15.4 | 18.8 | |
aThree control plots (Nos. 7, 10, and 13) were void of seedlings. | |||||
The loss of seedlings established by natural seedfall in plots numbered 14 to 41 is summarized in Table 13 (including death by deer trampling and digging by squirrels). Table 14 shows loss in one 0.6 x 12 meter (2 x 40 ft) burn trough which contained 2101 seedlings in a dense stand. Total losses to geometrids and gryllacridids was 23.7% at midsummer and increased slightly to 29.2% by fall. Both percentages are for the total number of seedlings present at that date. No seedlings were observed in control plots outside of the burn area.
Table 13. Random seedling study plots showing insect and other damage to natural regeneration in fuel reduction burn on Redwood Mountain—September 1970a.
| No. seedlings | No damage |
Hypocotyl. severed | 3 cotyledons eaten |
2 cotyledons eaten | 1 cotyledons eaten |
Maj. leaf dam. | Mod. leaf dam. |
Minor leaf coty. dam. | Seedlings with aphidsb |
Dead by deer | Dead by rodents |
Dead by drought etc. |
| 280c | 76 | 34 | 7 | 15 | 12 | 26 | 41 | 105 | 21 | 10 | 54 | |
| % of totala | 27.1 | 12.1 | 2.5 | 5.4 | 4.3 | 5.7 | 9.3 | 14.6 | 37.5 | 7.5 | 3.6 | 19.3 |
aSee appendix for complete account of each plot. bAlso counted in other categories. cTwenty-eight plots of 10 each. dTotal above 100% because of multiple conditions of seedlings. | ||||||||||||
Table 14. Insect loss to unusually dense seedlings in burned log trough (2101 individuals in 2 x 40 foot area) Redwood Mountain, Summer 1970.
Seedling Damage | ||||||||||||||||
| Portion of trough | No damage |
Hypocotyl severed | 3 cotyledons eaten |
2 cotyledons eaten | 1 cotyledons eaten |
Major leaf damage | Moderate leaf damage |
Minor leaf damage | ||||||||
| July | Sept | July | Sept | July | Sept | July | Sept | July | Sept | July | Sept | July | Sept | July | Sept | |
| West quarter | 208 | 101 | 69 | 11 | 18 | 0 | 17 | 0 | 18 | 0 | 19 | 30 | 13 | 5 | 53 | 4 |
| West mid-quarter | 398 | 357 | 34 | 9 | 11 | 1 | 16 | 1 | 18 | 0 | 14 | 66 | 2 | 4 | 49 | 4 |
| East mid-quarter | 330 | 184 | 4 | 19 | 2 | 0 | 7 | 1 | 2 | 1 | 26 | 36 | 0 | 6 | 22 | 17 |
| East quarter | 667 | 324 | 19 | 58 | 5 | 4 | 9 | 1 | 4 | 0 | 17 | 99 | 10 | 1 | 20 | 11 |
| Total | 1603 | 966 | 126 | 97 | 36 | 5 | 49 | 13 | 42 | 1 | 76 | 231 | 25 | 16 | 144 | 36 |
| % of total present at count | 76.3 | 70.8 | 6.0 | 7.1 | 1.7 | 0.4 | 2.3 | 0.9 | 2.0 | 0.1 | 3.6 | 16.9 | 1.2 | 1.2 | 6.9 | 2.6 |
Insect activity in cones
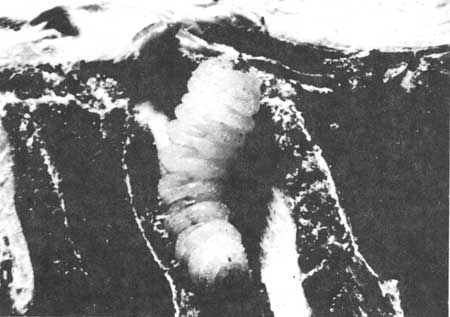
Oviposition by Phymatodes nitidus usually takes place in the cone peduncle attachment area. Larval stages usually penetrated and mined the axial shaft of the cones, reaching into individual cones scales (Fig. 50). Maximum projected linear length of mines was 21.2 cm. entering up to 15 scales. Larval frass was characterized by its fine, bicolored appearance. The particles which were tightly packed in the larval gallery resembled a coarse mixture of salt and pepper, composed of cream to white pieces mixed with amber to brown components. Frass was usually expelled in the terminal 5 to 12mm of gallery where overwintering pupation takes place. The diameter of these mature larval mines was less than 2 mm in width. Larval mining severed the vascular connections to the individual cone scales causing drying, shrinkage and the resultant release of the seeds that were borne on each scale. The resultant drying caused the browning of cones. Larval mining, exclusive of the cone, but still causing browning, occurred in the peduncles of 7% of the cones while peduncle mining in conjunction with that of the cone occurred in 13% of the brown cones sampled. Thus 80% of brown cones were mined in the body of the cone.
|
| Fig. 50. Larva of Phymatodes nitidus and mining activity in a giant sequoia cone. |
Giant sequoia cones remain alive up to 22 years (Hartesveldt et al. 1975). Unless severed from the tree by outside forces such as this beetle, they continue to add tiny annulations to the girth of the peduncle each year.
Adult Phymatodes emerge from the cones during early spring and summer, often before the snow has cleared from the groves. Emergence holes may appear anywhere on the cone, but most of them were found on the apical half. The holes were 1.0 to 1.5 mm in diameter and occurred on the flat apophysis of the scales. An emergence hole is illustrated in Fig. 51. Larval density averaged 1.4 individuals per cone in the Castro Tree. A maximum of 8 emergence holes in one cone were found in a tree growing at 2712 meters (8900 ft) in Sequoia National Park. In this tree, at the highest elevation known for the giant sequoia, the emergence holes averaged 4.6 per brown cone, three times greater than the majority of the trees which grow at lower elevations.

|
| Fig. 51. Giant sequoia cone with emergence hole of Phymatodes nitidus. |
The cone count made in August 1970 in the Castro Tree (Table 15) shows the vertical location and incidence of Phymatodes activity in cones based on external damage such as emergence holes and the presence of brown cone scales. Experience in rating damage has shown that an error of as much as 1.4% was possible in detecting the early stages of Phymatodes mining, even if external counts are made late in the summer.
Table 15. Distribution of ovulate cones and Phymatodes nitidus activity in the 290 foot Castro Tree (1970).
| Vertical Zones (ft) |
Total no. cones per level |
P.n.a cones per levelb |
Average % P.n. damage |
| 290-280 | 5212 | 856 | 16.4 |
| 280-270 | 4778 | 1031 | 21.6 |
| 270-260 | 3290 | 654 | 19.9 |
| 260-250 | 5777 | 1113 | 19.3 |
| 250-240 | 5361 | 1349 | 25.2 |
| 240-230 | 3231 | 583 | 18.0 |
| 230-220 | 2136 | 365 | 17.1 |
| 220-210 | 1705 | 293 | 17.2 |
| 210-200 | 2698 | 387 | 14.4 |
| 200-190 | 1089 | 113 | 10.4 |
| 180-170 | 560 | 37 | 6.6 |
| 170-160 | 2363 | 116 | 4.9 |
| 160-150 | — | 0 | 0.0 |
| 150-140 | 316 | 10 | 3.2 |
| 140-130 | 31 | 4 | 12.9 |
| 130-120 | 175 | 7 | 4.0 |
| 120-110 | 29 | 1 | 3.4 |
| 110-100 | — | 0 | 0.0 |
| 100-90 | — | 0 | 0.0 |
| 90-80 | 2 | 0 | 0.0 |
| 80-70 | 10 | 0 | 0.0 |
| Total no. cones | 39,508c | 6,964 | 17.6 |
aP.n = Phymatodes nitidus bIncludes brown cones (killed in past by Phymatodes nitidus, less proportioned browned by Gelechia (1.2%) and current season's beetle activity involving 1,016 cones in age classes over 2 years. c20,697 cones are new cones (1970 had an unusually large crop). | |||
Following the heavy cone crop of 1970 the Castro Tree held a total of 39,508 cones, 70% of which were located in the upper third of the tree. There were 5,948 brown and 20,697 first year cones. Cones of age classes two to twenty years and older numbered 12,836.
A total of 3,147 brown and semibrown cones were dissected by the double axial-slice technique. Phymatodes nitidus mining was found in all age classes examined (Table 16). The cones were selected from the total available in the Castro Tree by self-weighted, stratified random sampling. Over 8.4% of the attacks occurred in age classes of cones four through nine years. Sixteen brown cones did not show any insect mining and may be representative of reproductive tissue lost to weather factors, or quite possibly were damaged by the researcher within the tree. The bulk of the cones (34,188 or 86.5%) as well as the Phymatodes activity (6631 or 95.2%) occurred in the top half of the foliated portion of the Castro Tree at heights of 60 to 88 m (200-290 ft). A total of 302 cones out of the 3,147 sampled were mined by both Phymatodes and a gelechiid moth, Gelechia sp.
Table 16. Age classes of 3,147 giant sequoia cones attacked by Phymatodes nitidus in the Castro Tree (August 1969).
| Age class of conesa |
Cones with P.n.b |
Percent of total |
Age class of cones |
Cones with P.n. |
Percent of total | |
| 1 | 0 | 0.00 | 11 | 54 | 1.72 | |
| 2 | 51 | 1.62 | 12 | 81 | 2.57 | |
| 3 | 64 | 2.03 | 13 | 74 | 2.35 | |
| 4 | 308 | 9.79 | 14 | 15 | 0.48 | |
| 5 | 373 | 11.85 | 15 | 3 | 0.10 | |
| 6 | 541 | 17.19 | 16 | 2 | 0.06 | |
| 7 | 507 | 16.11 | 17 | 8 | 0.25 | |
| 8 | 424 | 13.47 | 18 | 1 | 0.03 | |
| 9 | 514 | 16.33 | 19 | 0 | 0.00 | |
| 10 | 122 | 3.87 | 20 | 2 | 0.06 | |
aAge class in years bP.n. stands for Phymatodes nitidus | ||||||
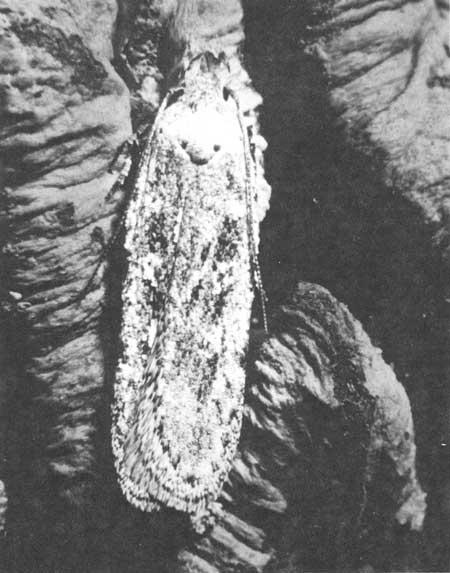
During the summer of 1968 small gelechiid larvae were noticed boring into giant sequoia cones. They were identified as Gelechia sp. by R. W. Hodges of the U.S. National Museum. Unlike Phymatodes nitidus, this moth attacks only first year cones and attains the adult stage only about 5% of the time (Fig. 52).
|
| Fig. 52. Adult of Gelechia sp. |
Oviposition by Gelechia occurs in first year cones between late June and early August, and takes place at the outside juncture of adjoining cone scales. The fleshy first year cones are relatively soft and apparently provide a feeding substrate for the developing larvae. Resinous cone fluid was exuded at the larval entrance sites on the outer surface of the cone and often trapped early instars of the moth. A three year survey of incidence of new attacks on cones by the moth in the Castro Tree showed a range of from 2 to 27% damage. Differences varied with location on the tree and with the season. A possible correlation between size of cone crop and rate of larval intrusion was illustrated by the fact that in 1968 and 1969, when first year cones occurred in average numbers, 2.6 and 3.5% of the new cones were attacked. In 1970 when the new cone crop was extremely large, the total moth attack (of new sequoia cones) was 27.3%. Other conifers of Redwood Canyon had similar increase in cone yield in 1970. Table 17 shows the relationship of the 1970 cones attacked by Gelechia to elevation in the Castro Tree. Gelechiid incidence was distinctly higher in the upper crown of the tree.
Table 17. Distribution of first year ovulate cones and Gelechia sp. activity in the 88 m (290 ft) Castro Tree (1970).
| Vertical Zones (ft) |
Total no. cones per level |
Gelechia cones per level |
Average % Cone attack |
| 290-280 | 3215 | 1531 | 47.6 |
| 280-270 | 3168 | 1349 | 42.6 |
| 270-260 | 1387 | 413 | 29.8 |
| 260-250 | 2921 | 890 | 30.5 |
| 250-240 | 2880 | 627 | 21.8 |
| 240-230 | 1567 | 197 | 12.6 |
| 230-220 | 1429 | 289 | 20.2 |
| 220-210 | 960 | 121 | 12.6 |
| 210-200 | 1025 | 174 | 17.0 |
| 200-190 | 306 | 41 | 13.4 |
| 190-180 | 483 | 22 | 4.6 |
| 180-170 | 203 | 6 | 3.0 |
| 170-160 | 1064 | 98 | 9.2 |
| 160-150 | 0 | 0 | 0.0 |
| 150-140 | 66 | 2 | 3.0 |
| 140-130 | 1 | 0 | 0.0 |
| 130-120 | 21 | 0 | 0.0 |
| Total no. cones | 20,697 | 5,649 | 27.3 |
The survival rate of cone feeding Gelechia larvae was only slightly higher in low cone yield years than in the extremely heavy cone crop year of 1970. The maturation of larvae was 4.3% in cones in 1969 versus 4.9% in 1970. The number of attacks per cone was easy to distinguish because of the resinous exudate at each entrance point. A mean of 1.7 Gelechia entrances per new cone were recorded in the Castro Tree in 1970 with a maximum of five entrances per cone.
Larval mining by this moth differed from that of Phymatodes by the presence of silk in the frass, point of entry and the tendency of Gelechia to feed within 3 to 4 scales in a group rather than to feed lengthwise along the cone axis. Cones first thought to be genetic "doubles" were, on investigation, found to be caused by atypical longitudinal mining in the axial shaft by Gelechia. In these cases the shaft split into two lengthwise halves bowed apart in the center, thus giving the double cone effect (Fig. 53). Two samples of 60 and 100 "double cones," when cut open, all showed gelechiid larval damage. None of the several hundred cones observed were completely brown from Gelechia activity, with the exception of several undersized defective cones. Gelechia may be very numerous in younger sequoia (150-200 ft class). Forty-seven percent of the cones in trees of this size studied in 1974 showed past or present Gelechia damage.

|
| Fig. 53. Double cone or "twin cones" of giant sequoia induced by gelechiid moth infestation. |
Early instars of a small lygeid bug were noted feeding upon seeds in newly opened cones. These insects have sucking mouth parts which remove fluid from seeds that have not been dislodged from the open cones. Later growth stages, including the adults of this insect, Ischnorrhynchus resedae, retreat deep within the open cone scales when not feeding upon foliage of adjacent new terminals.
Discussion and summary
Seedling damage by insects
The differences in observed mortalities through time and between plots can be explained on the basis that those areas experiencing recent disturbances such as burning and/or manipulation appeared to sustain greater insect damage. This parallel between extent of insect damage and a severe disturbance, such as a fire, relates to the earlier observation that under those conditions sequoia seedlings were about the only source of food. Later, as more plants become established, general foliage feeding insects such as the ones reported in this study return to other more desirable plants. The data agree with this hypothesis. Plot 1, established in an eight acre 1965 burn area, showed a significantly greater loss compared to the loss in plots 2 and 3 which were established in an undisturbed forest. The two transects illustrated the same trend where South Area, which burned in 1966 or one year later than Trail, indicated slightly higher seedling loss.
After observing these differences, the decision was made to more closely monitor transects and massed seed beds during the summers of 1969 and 1970. Information was needed to answer questions as to the degree of damage to seedlings short of death, density and differences in damage to seedlings in fresh burn areas versus older and nonburn plots, and the time of year that damage occurred. The 17.5% mortality in the new burn seedling plots (4-13) was almost triple that of prepared seed beds in areas burned two years previously. Plots 14 to 41, randomly selected from within the 1000 foot wide 1970 NPS environmental burn, registered 12.1% mortality due to insects excluding other lesser damage that likely caused mortality at a later date. Field checks of these plots in late summer of 1971 showed loss of 3.4% seedlings to that year's insect activity. Again these data suggest that the largest loss of seedlings came immediately after a burn.
Release of seed from sequoia cones
The release of seed initiated by Phymatodes nitidus activity in the ovulate cones of the giant sequoia is of utmost importance from the standpoint of timing and point of release of seed from the tree. Cones damaged by this beetle will open after drying during the late summer and fall months, which, in effect, may lessen the exposure of the seed to lethal radiation and desiccation. Damaged cones will open several scales at a time over a period of six months to a year. Seed drop was also reported throughout the winter as evidenced by fresh fall on snow (Benner pers. comm.)
It was observed as early as 1967 that seeds falling from the 250 foot level of the Albina Tree would be displaced several dozen meters off the vertical fall line, even on windless days. Winds that accompany late summer Sierran storms strike Redwood Canyon at velocities of from 48 to 64 kilometers (30 to 40 miles) per hour or greater. These storms coincide with fall seed drop from brown cones and cover the forest floor with seeds after every heavy wind. Seeds dropped at this time could be disseminated for at least a quarter of a mile downwind from the parent tree. For example, field tests in the Castro Tree (1969, 1970) showed seeds fall an average 1.8 m (6 ft) per second. Theoretically a wind of 64 kph (40 mph) across the top of the sequoia could move seed 291 m (957 ft) in the first 16.5 seconds in a drop from 8.5 to 55 m (28.0 to 180 ft), which is from sequoia crown to the top of the lower canopy of fir, pine and cedar. Additional horizontal movement takes place through this lower canopy at greatly reduced wind speeds. If the wind velocity averages 24 kph (15 mph) (a very low estimate) in a drop from 55 m (180 ft) to the ground, the seed could travel another 210 m (690 ft) in 30 seconds of descent, or a total dispersal of 502 m (1647 ft) lateral displacement. This dispersal of well over 0.4 kilometers (one-quarter mile) need only occur once in every 100 years or more for sufficient reproduction but probably happens several times a summer and with even greater frequency in winter.
Measurement of vertical and lateral seed motion in a uniform airstream is acceptable only in gaining averages for seed dispersal in the many variable sites and tree configurations. Factors such as gustiness and updrafts caused by cover turbulence affect vertical range of drop (Kittredge 1948).
Relationship and role of cone insects to other reproductive forces
The two principle cone insects avoid competition with each other by feeding on different year classes. Phymatodes nitidus mainly mines cones of age classes four years and older while the cone boring moth, Gelechia, feeds only upon the first year cones (damage may cause some seeds to be released through the second or third year). Such a relationship of sequential feeding is not unique. Coulson and Franklin (1968) found similar selective behavior with Dioryctria coneworms and cecidomyiid cone maggots, which attacked 14.3% of the second year cones of the shortleaf pine, Pinus echinata. They noted that fresh first-year cones were not mined by Dioryctria, while dried first-year cones were fed upon by the gelechiid, Battaristis vitella. The latter could not be induced to feed on living cones.
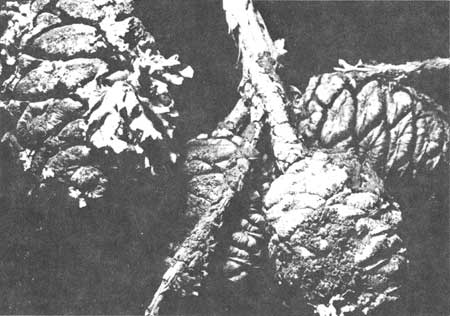
Lichens begin to grow on the exterior surface of the sequoia cones at about the time that Phymatodes nitidus activity and the optimal seed viability of the giant sequoia diminishes. Such growth may prevent the seeds from falling free of the cones (Fig. 54). Two species of lichens may completely cover individual cones or, in some cases, completely conceal entire cone masses. The predominant lichens growing on cones in the age classes beyond eight years are Parmelia physodes, a foliose form; Letharia vulpina, a chartreuse dendritic form; and Hypogymina imshangii, a black and white inflated lobe type. Small specks of lichen growth usually appear prior to eight years but probably do not impede seed drop.
|
| Fig. 54. Lichen covered cones of a giant sequoia. |
Several physical factors may aid in cone release and subsequent dispersal. The more important factors include snow loading of branches, ice deposits from ice storms, and heavy winds which may cause an increase in the drop of cones and may be of sufficient magnitude to release many seeds possessing strong attachment to the open cones. Seeds of this type examined from the top of the Castro Tree in 1969 and 1970 showed that 75% (by snap test) were viable, which is unusually high for sequoia seed (Chapter 5).
Fire is the most important physical agent concerned in reproduction of the giant sequoia, as was discussed in Chapter 5. Super-heated air may cause death of the cones with resultant release of large quantities of seed.
In summary, insects rarely attacked the 1-2 year old seedlings of giant sequoias except where they grew in dense numbers. The cone insects are also few in number, namely a moth that damages first year cones and a minute beetle that causes cones to drop their seeds. The latter insect is very important in the continual release of seeds and subsequent reproduction of the giant sequoia.
| <<< Previous | <<< Contents >>> | Next >>> |
chap7.htm
Last Updated: 06-Mar-2007