|
NATIONAL PARK SERVICE
Giant Sequoia Ecology: Fire and Reproduction |

|
CHAPTER 8:
BIRDS AND MAMMALS, FIRE, AND GIANT SEQUOIA REPRODUCTION
Howard S. Shellhammer
Introduction
Giant sequoia groves do not possess a special vertebrate fauna. The fauna of the groves varies with altitude and latitude, just as it does in the mixed coniferous forest that surrounds them and which runs the length of the Sierra Nevada.
The mammals of the Redwood Mountain Grove are dominated by rodents: deer mice (Peromyscus maniculatus), Douglas squirrels (Tamiasciurus douglasi) and to some degree by California ground squirrels (Otospermophilus beecheyi) and chipmunks (Eutamias spp.). A listing of the other mammals, amphibians and reptiles, all common to much of the Sierra Nevada, can be found in Hartesveldt et al. (1975). Wallis (1951) reported on vertebrates in the Mariposa Grove. An extensive list of the breeding birds in one portion of the Redwood Mountain Grove was compiled by Kilgore (1971a) and is applicable to our present study areas. Dominant species include the dark-eyed junco (Junco oreganus), mountain chickadee (Parus gambeli), robin (Turdus migratorius), black-headed grosbeak (Pheucticus melanocephalus), western tanager (Piranga ludoviciana), western wood peewee (Contopus sordidulus), and red-breasted nuthatch (Sitta canadensis).
Few studies have been carried out to determine the relationship between fire and the birds and mammals of the groves. Kilgore (1971) studied the changes in the numbers of breeding birds in Whitaker's Forest following manipulations similar to the present study. Whitaker's Forest, a 130 hectare (320 acre) experimental forest of the University of California on the western slope of Redwood Mountain, is located within the boundaries of the Redwood Mountain Grove (Fig. 3).
Little information other than conjecture has been made as to which birds and mammals interact directly with the giant sequoia or affect its reproduction. Ingles (1965) and Fry and White (1930) noted that Douglas squirrels cut and cached giant sequoia cones and also inferred that they consume their seeds. Fry and White (1930) presented no figures but gave the following description of seedling—animal interactions:
Among the conifers I know of no other that is attacked in infancy by so many destructive agencies as the Big Tree. No sooner are the seedlings above the ground than destruction and disaster assail them, so that few survive the first year. Birds, such as sparrows and finches, are attracted to the tender green plants with their seed hulls on their tops and pick off the topmost portion for food. This invariably kills the plants. Cutworms destroy many of the plants by cutting them off near the ground. Large blackwood ants levy a heavy toll by cutting off the plants and dragging them to their nests. Rodents, such as ground squirrels and chipmunks, destroy many of the plants by eating off the tender tops.
The acceptance of some of these generalizations must be tempered by the fact that many of the observations were made in nursery conditions outside of a grove, where seedling sequoias were being grown in thick beds.
The present study attempts to ascertain whether the vertebrate interactions described by Fry and White (1930) are correct and to gain some idea of the magnitude of the effect of birds and mammals on sequoia reproduction. It also attempts to identify the changes taking place in the number of birds and mammals following the use of fire and associated manipulations in a grove. This study dwells upon the impact of birds and mammals on the seed and seedling stages of the giant sequoia.
Methods
Most of the studies reported on in this section took place in Ridge, North and Trail Areas (Chapters 2 and 3 for descriptions, Fig. 4). Miscellaneous investigations took place in and near the Redwood Mountain and General Grant Groves of Kings Canyon National Park and the Giant Forest of Sequoia National Park (Fig. 3).
Avian studies
Populations of some of the species of breeding birds, in both the manipulated and control portions of Ridge, North and Trail Areas were measured in 1965 and 1966 by the plot census method (Kendeigh 1944), using dittoed maps on which to record observations. This same method was used by Kilgore (1971a) in studying bird populations on the other side of Redwood Mountain during the same years. Routes were taken through the study areas such that the observer came within 30.5 m (100 ft) of every point in each area. Activities of all birds were recorded, with special emphasis given to singing males. Five censuses were made between late June and early August in 1965 and again in 1966. Approximate territories or areas of activity were identified at the end of the season by circling groups of observations on the dittoed maps.
Mammalian studies
Small mammals were trapped using mouse and rat-sized Sherman live traps in both the manipulated and control portions of North Area in 1964, and in Ridge, North and Trail Areas in 1965, 1966 and 1967 for a total of 5527 trap nights (Fig. 55). Captured mice and chipmunks were tagged with numbered ear tags, measured, checked for sexual condition, and released. Each area was trapped between five and eight times per summer. The density of mice per area was calculated using the technique developed by Brant (1962) of adding a border to the trapping area equal to the average distance moved of the animals in that population for that particular trapping period.

|
| Fig. 55. Live trap held by Howard Shellhammer. |
Seedling studies
Chipmunks, ground squirrels and Douglas squirrels were censused using the plot census method developed for birds by Kendeigh (1944). The method was modified to include long periods of direct observation from fixed points. None of these larger rodents were ear-tagged. Chipmunks and ground squirrels were censused from 1965 through 1967 and Douglas squirrels from 1965 through 1967 (Chapter 9) for a total of approximately 650 hours of observation.
Deer and other large mammals, such as coyotes and grey foxes, were observed, or the evidence of their activity was noted in the censusing mentioned previously or during the many hundreds of other hours spent in or near the study areas.
Seed preference studies
Seed preferences were checked using a modification of the seed spot method which Tevis (1956) modified from Moore (1949-50). Seeds of seven conifers, as well as sunflower, were buried in 1.3 cm (0.5 in) deep pits spaced 60 cm (24 in) apart in cleared areas on the forest floor. Generally eight pits, one of each kind of seed, were used in each experiment, which yielded 4.5 meter by 60 cm cleared areas. Fifteen of each type of seed, except giant sequoia seeds where 45-50 were used, were buried in each pit. The sequence of seeds (pits) in an experimental area was randomly rearranged if more than one series of tests were run in an area during the summer. Thirty-five series (278 pits) were run in the Redwood Mountain Grove in 1965, as were 31 series (249 pits) in 1966. Ten series (80 pits) were run in a non-sequoian, mixed conifer forest near the Grant Grove in 1965. Conifer seeds included sugar pine (Pinus lambertiana), ponderosa pine (P. ponderosa). Jeffrey pine (P. jeffreyi), white fir (Abies concolor), red fir (A. magnifica), incense cedar (Calocedrus decurrens), and giant sequoia. Jeffrey pine and red fir were not found in the test areas. All conifer seeds used were collected by the California Division of Forestry from areas close to the Redwood Mountain Grove. The activities of various species of rodents were easily discernible. Deer mice dug simple, neat pits over the seed spots while chipmunks and Douglas squirrels undertook considerable exploratory digging and made larger pits. Birds did not uncover seeds buried in this manner.
All seedlings germinating and observable in the manipulated areas were marked and observed for animal damage during the period of the study or until they died (Chapter 5 for seedling mortality studies). Stecker (Chapter 6) estimated the loss of seedlings to insects in various plots and in transects both in and outside of the study areas.
Seedlings of giant sequoias, incense cedars, Jeffrey pines, and red firs were grown in 40.6 x 20.3 x 15.2 cm boxes in a greenhouse at San Jose State University in San Jose, California, and the boxes were placed in the treated and control portions of North and Trail Areas in early July 1966. Observations were made on the fate of 174 seedlings ranging from 1-2 cm giant sequoias to 7-19 cm red firs from July 4 to August 15, 1966.
Results
Avian studies
The territories of only seven species of birds, plus the empidonax complex of flycatchers, could be assessed with certainty due to the lateness of the observation periods in both years (Table 18). Empidonax flycatchers increased slightly in the control plots and decreased in the manipulated plots in North Area. Mountain chickadees increased in both plots in Ridge but not in North Area. Dark-eyed juncos showed only a slight decrease in treated versus control plots, whereas western wood peewees increased in the treated areas. A noticeable, but unsubstantial, increase in the number of robins occurred in the treated areas. These results were similar to those obtained by Kilgore (1971a) in 18.2 hectare (40 acre) manipulated and control plots in Whitaker's Forest during the same years (1965 and 1966). Both studies showed similar changes in empidonax flycatchers, mountain chickadees, western wood peewees, and robins, while Kilgore noted a larger decrease in dark-eyed juncos in treated plots. Relatively small and similar changes in avifauna were observed in our small manipulated areas (approximately 2 hectares) and the 18.2 hectare areas studied by Kilgore. The type of habitat manipulations in both studies were generally similar.
Table 18. Numbers of pairs of birds per hectare in Ridge, North and Trail Areas before and after manipulation and burning.a
| Ridge Area |
North Area |
Trail Area | ||||||||
| Feeding height and species |
Control | Burn | Control | Burnb | Preburn | Postburn | ||||
| 1965 | 1966 | 1965 | 1966 | 1965 | 1966 | |||||
| UPPER CANOPY | ||||||||||
| Empidonax sp. | 0 | 0 | 0 | 0 | 2 | 3 | 1.5 | 0.5 | 0.5 | 0.5 |
| Western tanager | 1 | 1 | 1 | 1 | 1.5 | 1 | 1 | 1 | 0.8 | 0.5 |
| UNDERSTORY | ||||||||||
| Western wood peewee | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 2 | 0.5 | 0.5 |
| Mountain chickadee | 2 | 3 | 1 | 3 | 1 | 1 | 1 | 1 | 0.5 | 1 |
| BRUSH/SAPLING | ||||||||||
| Winter wren | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0.5 | 1 |
| Townsend's solitaire | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.5 | 0 |
| TRUNK GROUND | ||||||||||
| Hermit thrush | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0.3 | 0.5 |
| Black-eyed junco | 2 | 1.5 | 1 | 1 | 4 | 5 | 4 | 4 | 1.5 | 1.0 |
aRidge and North Areas had controls while Trail Area did not. b1965burn area in North Area was partially manipulated in the fall of 1964 and was burnt again in the fall of 1965, hence 1965 is realty postmanipulation #1 and 1966 is postmanipulation #2. All areas listed are about 1 hectare except Trail which was approximately 2 hectares. | ||||||||||
Apparent sapsucker damage was observed on the upper portions of nearly all the larger giant sequoias inspected (Fig. 56). Such damage is attributed to sapsuckers because of the characteristic pattern of holes in the thinner portions of the bark, even though no researchers, past or present, have seen sapsuckers working the trees. A survey of 32 trees in and near South Area showed that all but two of the 26 trees with a diameter at breast height (dbh) of 1.8 m (4.5 ft) or larger had moderate to extensive drilling damage on the upper two-thirds of their trunks. Much of this damage appeared to be old, but no estimate could be made of how old. Trees with a dbh of less than 1.8 m (4.5 ft) seldom showed such damage. A similar situation existed in the other study areas.

|
| Fig. 56. Apparent sapsucker damage on the trunk of a giant sequoia. |
Stecker (Chapter 7) noted sapsucker damage near the top of the large giant sequoia in which he worked in 1968. He noted that it was extensive enough to impair vascular transport and apparently was resulting in the retardation of growth of the leader shoot of the tree.
Mammalian studies
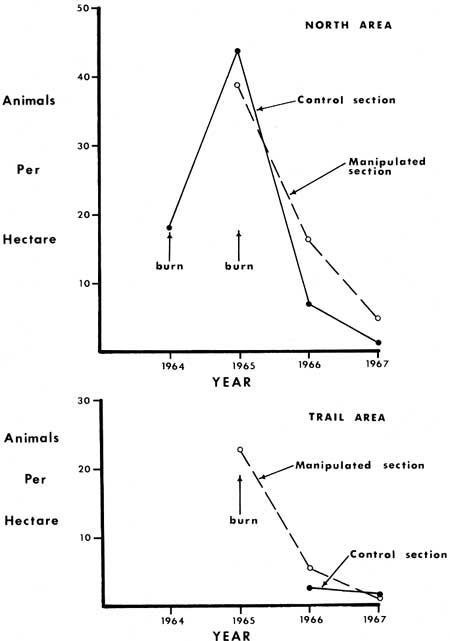
Deer mice (Peromyscus maniculatus) were the most common mammals in the study areas (Fig. 57). They were most numerous in 1965 but declined each year for the next two years (Table 19, Fig. 58). Approximately two times as many mice were trapped in the treated versus control portions of North Area in 1966 and 1967 and Trail Area in 1966. There were, however, slightly more mice in the control portion of North Area in 1965, a fact possibly associated with the sequence of manipulations of that area. North Area was first manipulated and partially burned in the fall of 1964, but the fire did not carry well due to the mechanical disturbance of the litter surface. The piles of logs were reburned in the fall of 1965.
|
| Fig. 57. A deer mouse, Peromyscus maniculatus, tagged in the ear with a light-weight, numbered tag. |
Table 19. Numbers of deer mice, Peromyscus maniculatus, in North and Trail Areas before and after manipulations.
| Year | Animals captured twice or more |
Ajusted area in hectaresa |
Density in hectares |
Av. D.b in meters |
| North Area CONTROL | ||||
| 1964 | 20 | 1.1 | 18.0 | 21.5 |
| 1965 | 34 | 0.8 | 43.7 | 13.7 |
| 1966 | 23 | 3.4 | 6.7 | 30.7 |
| 1967 | 6.5 | 4.8 | 1.2 | 69.6 |
North Area MANIPULATED | ||||
| 1964 | — | — | — | — |
| Burnt in fall of 1964 | ||||
| 1965 | 36 | 0.9 | 38.7 | 13.7 |
| Piles reburnt in fall of 1965 | ||||
| 1966 | 15 | 0.9 | 16.1 | 30.7 |
| 1967 | 4.5 | 0.9 | 4.9 | 73.8 |
Trail Area CONTROL | ||||
| 1964 | — | — | — | — |
| 1965 | — | — | — | — |
| 1966 | 4 | 1.7 | 2.5 | 27.6 |
| 1967 | 5 | 3.0 | 1.7 | 58.6 |
Trail Area MANIPULATED | ||||
| 1964 | — | — | — | — |
| 1965 | 45 | 1.9 | 22.7 | 14.2 |
| Burnt in fall of 1965 | ||||
| 1966 | 15 | 2.7 | 5.4 | 27.6 |
| 1967 | 5 | 4.3 | 1.2 | 56.7 |
aArea is adjusted by adding a band equal to the average distance moved between captures of animals in that population. bAv. D. is average distance moved between captures. | ||||
|
| Fig. 58. Densities of Peromyscus maniculatus in North and Trail Areas. |
The reproductive activity of these mice was high in 1965 when an estimated 184 young were added to North Area during the time of the summer study period. Fewer of these young animals were trapped than expected. There was no evidence of emigration, as there was in 1966 in the same area. One male of the smaller 1966 population moved 400 meters (1,320 ft) down the canyon to near Trail Area. The implications of such a move in relationship to emigration is grossly conjectural because of the single observation. In addition, the average distance moved between captures, Av. D, (Table 19), was inversely related to density, with the Av. D. being the smallest in 1965. Predators such as coyotes and owls were associated with the areas, however the coyote scats examined contained few remains of mice. The effects of the owls and other birds of prey on mouse populations were not considered in this study.
The squirrels and chipmunks present included the grey squirrel (Sciurus griseus), Lodgepole chipmunk (Eutamias speciosus) and Townsend's chipmunk (E. townsendii) in addition to the Douglas squirrels and California ground squirrels mentioned earlier. The numbers of the other species increased in 1966 and dropped in 1967 (similar to, but not as dramatically as did the Douglas squirrels). There was little difference in their numbers between the treated and control portion of the study areas, although such distinctions were made difficult by the relatively small sizes of the plots.
A grey squirrel inhabited part of North Area in 1965 and 1966, and another was observed in Trail Area from 1965 to 1967, as well as in later years of the study, when squirrels other than the Douglas squirrel were receiving little attention. One dug up two sequoia cones in Trail Area in 1966 and ate on them in a manner similar to that used by Douglas squirrels (Chapter 9). The cones were partially trimmed by this animal, and one of them was reburied. It is doubtful that these cones were previously cached by Douglas squirrels because the site was too open and the soil was too dry. Grey squirrels ate sugar pine cones, often just before the time of the year that Douglas squirrels started to cut them. This was especially true in the late summer of 1974 when there were few Douglas squirrels, and most conifers, excluding giant sequoias, had very few cones on them. Several groups of sugar pines which had moderately good cone loads were stripped of their cones by grey squirrels, and the cones were then eaten near the bases of the trees. No instances were observed of grey squirrels cutting or caching giant sequoia cones except for the instance previously mentioned. Grey squirrels may have eaten some fallen sequoia cones, but the number was insignificant compared to the numbers cut by Douglas squirrels (Chapter 9).
A California ground squirrel was observed to eat part of a Jeffrey cone at a site two kilometers upslope from Ridge Area, but that was the only incidence of that species feeding on cones during the term of the study. No observations were made of this species eating any giant sequoia materials.
Trowbridge shrews (Sorex trowbridgii) were present in North and Trail Areas in low numbers (1 to 4 animals) in 1965 and 1966. One mole (Scapanus latimanus) was known to be present in North Area in 1966. While their diggings were not seen, pocket gophers (Thomomys bottae) were observed in the talons of goshawks as they moved through North and Trail Areas on several occasions in 1965 and 1966. One meadow mouse (Microtus loingicaudus) was captured in North Area in 1965, and two were captured in 1966. Several small white firs in the control portion of North Area were partially girdled (one killed) by meadow mice in the winter or early spring of 1965-66. A meadow mouse was observed in the same area in the summer of 1966 eating the tips of small white firs. No damage positively attributable to meadow mice was observed on any giant sequoia.
Seed preference studies
Giant sequoias shed enormous numbers of seeds. My best estimate is 180 seeds/m2/yr under mature trees while Harvey (Chapter 5) noted a variation on the ground of a few dozen to several thousand seeds/m2/yr. The former figure was arrived at by multiplying the number of seeds per cone (200) times the average number of cones shedding seeds per tree per year (1,500) times the number of mature trees per hectare (6) divided by 10,000 (i.e. m2/ha). Such shed seeds were considered initially to be a potential food source for the animals of the grove.
Seed preferences of various rodents, as tested in the seed spot tests, are listed in priority order on Tables 20 and 21. Chipmunks, and possibly a few Douglas squirrels, accounted for 60% of the seed spots uncovered in 1965 and 95% of those uncovered in 1966. Deer mice accounted for the difference each year. The variation between years is correlated with rodent densities. Deer mice were four times more plentiful in 1965 in trapping grids near the seed spot test sites than in 1966 (Table 19). Chipmunks and Douglas squirrels were both twice as numerous in 1966 as in 1965 (Chapter 9, Table 24).
Large pine seeds were the preferred type of food each year, regardless of whether they were native to the area of the test or not. Sunflower seeds were also highly desired. They were successfully used as bait in trapping small mammals in areas away from the seed preference sites. Giant sequoia seeds were the least preferred, apparently because they were the smallest in size and weight, and were partially eaten in only 14% of the spots inside the grove and 19% of the spots outside in 1965 (Tables 20, 21). Eight of the nine spots partially eaten that year were eaten during the first run at any particular site. All of the spots of sequoia seeds were exposed but not eaten during the second run one week later at the site outside the grove, while approximately 50% of the spots inside the grove were not exposed at all, and only one was exposed and partially eaten. Ninety-four percent of the spots of giant sequoia seeds were uncovered but not eaten in 1966. Rodents generally preferred large seeds, although the highly preferred ponderosa pine seeds were relatively small (Table 22).
Table 20. Preference of rodents in 1965 seed spot tests at Redwood Mountain Grove and near Grant Grove.
| Rank | Seed | Percent exposed, eaten |
Percent exposed, partially eaten |
Percent exposed, not eaten |
Percent not exposed |
No. of tests |
| 1 | Sugar pine | 100.0 | — | — | — | 25 |
| 2 | Ponderosa pine | 100.0 | — | — | — | 25 |
| 3 | Jeffrey pine | 100.0 | — | — | — | 25 |
| 4 | Sunflower | 100.0 | — | — | — | 25 |
| 5 | White fir | 91.0 | 9.0 | — | — | 35 |
| 6 | Red fir | 72.0 | 28.0 | — | — | 25 |
| 7 | Incense cedar | 25.5 | 43.0 | 25.5 | 6.0 | 59 |
| 8 | Giant sequoia | — | 14.0 | 43.0 | 43.0 | 59 |
Near Grant Grovea | ||||||
| 1 | Sugar pine | 100.0 | — | — | — | 8 |
| 2 | Ponderosa pine | 100.0 | — | — | — | 8 |
| 3 | Jeffrey pine | 100.0 | — | — | — | 8 |
| 4 | Sunflower | 100.0 | — | — | — | 8 |
| 5 | Red fir | 100.0 | — | — | — | 8 |
| 6 | White fir | 88.0 | 12.0 | — | — | 8 |
| 7 | Incense cedar | 44.0 | 31.0 | 26.0 | — | 16 |
| 8 | Giant sequoia | — | 19.0 | 81.0 | — | 16 |
aOutside of Grant Grove in a mixed coniferous forest. | ||||||
Table 21. Preference of rodents in 1966 seed spot tests at Redwood Mountain Grove.
| Rank | Seed | Percent exposed, eaten |
Percent exposed, partially eaten |
Percent exposed, not eaten |
Percent not exposed |
No. of tests |
| 1 | Sugar pine | 96.6 | — | — | 3.4 | 29 |
| 2 | Ponderosa pine | 92.0 | — | — | 8.0 | 25 |
| 3 | Sunflower | 92.0 | — | — | 8.0 | 25 |
| 4 | Jeffrey pine | 88.0 | — | — | 12.0 | 25 |
| 5 | Red fir | 56.0 | 28.0 | 12.0 | 4.0 | 25 |
| 6 | White fir | 37.9 | 48.3 | 13.8 | — | 29 |
| 7 | Incense cedar | — | 15.8 | 71.1 | 13.2 | 38 |
| 8 | Giant sequoia | — | — | 94.3 | 5.7 | 53 |
Table 22. Seed preference of rodents versus average seed weight.
| Weight of seeds in grams | |||||||
| Rank | Seed | 0.20 | 0.15 | 0.10 | 0.05 | 0.01 | 0.005 |
| 1 | Sugar pine | 0.22 | |||||
| 2 | Ponderosa pine | 0.05 | |||||
| 3 | Sunflower | 0.10 | |||||
| 4 | Jeffrey pine | 0.12 | |||||
| 5 | Red fir | 0.09 | |||||
| 6 | White fir | 0.04 | |||||
| 7 | Incense cedar | 0.03 | |||||
| 8 | Giant sequoia | 0.005 | |||||
Seedling studies
Desiccation accounted for 84.6% of the mortality in 5206 marked seedlings (see seedling mortality studies, Chapter 5). Insects caused another 5.7%, while birds and mammals killed 1.1% of the total. Included in those killed by birds and mammals were three seedlings trampled by deer and 15 crushed by horses. No fatal damage caused by birds, mammals or insects was observed in seedlings older than three years (Chapters 5 and 7).
Stecker's studies outside the study areas (Chapter 7) revealed much the same pattern. He reported slight loses to deer browse in first year seedlings.
No damage to deer browse was noted on older giant sequoia seedlings or young trees, although Ceanothus parvifolius and other shrubs both inside and out of the study areas were browsed in various years. A few four and five year old giant sequoia seedlings were browsed by deer in an area manipulated by the park service on the ridge of Redwood Mountain.
Moderate, damage to boxed seedlings can be attributed to birds or mammals. Twelve of the 48 2-4 cm giant sequoias killed during the period of observation were killed either by insects or birds (Table 23). Insects are suspected in this case, as five other seedlings in a nearby box were chewed upon by insects but not killed. A tight bunch of five seedlings were crushed by a deer as it moved through the area of the boxes. Ten of the larger seedlings were dug up, and the box in which they were growing was partially displaced, presumably by California ground squirrels which had their den close by. None of the largest giant sequoia seedlings (4-6 cm, Table 23) were eaten by vertebrates, although some were dug up, as mentioned above. Thus twenty-five percent (32) of the 128 boxed sequoia seedlings were killed during the test, with the heaviest damage in the intermediate size group (2-4 cm).
Table 23. Damage to laboratory-grown seedlings placed in study areas.
| Species and size | N | Number damaged |
Type of damage |
| >Giant sequoia, 1-2 cm | 18 | 2 | 1 eaten by insects, 1 unearthed |
| Giant sequoia, 2-4 cm | 48 | 22 | 5 crushed by deer, 5 chewed on by insects but not killed, 12 topped and killed by insects or birds |
| Giant sequoia, 4-6 cm | 62 | 10 | 10 dug up by rodents |
| Incense cedar, 5-7 cm | 21 | 0 | — |
| Jeffrey pine, 12-14 cm | 15 | 0 | — |
| Red fir, 7-19 cm | 10 | 0 | — |
One pertinent direct observation was made in 1966 of the interaction of a female purple finch and several boxes of seedlings. The bird spent several minutes moving through the boxes pecking at the surface of the soil between seedlings. None of the seedlings were eaten by the finch.
Discussion
Effects of fire on birds and mammals
Experimental burns of two to four hectares produced relatively minor changes in bird and small mammal populations. These small burns resulted in decreases in empidonax flycatchers and dark-eyed juncos and increases in western wood peewees and robins. Similar results were observed by Kilgore (1971) in larger (18.2 hectare) burns. The dominant small rodent, Peromyscus maniculatus, was 2.1 to 2.4 times more numerous in burned areas than in control areas in the year following the burn. Similar increases after burns of various sizes have also been described by Alhgren (1966), Cook (1959), Garman and Orr-Ewing (1949), Gashwiler (1970), and Tevis (1956). The numbers of Douglas squirrels appeared to be affected by other factors than those resulting from fires (Chapter 9).
Bendell (1974) summarized numerous studies and observed that most populations (80%) of birds and mammals studied showed no significant changes after fires. He noted a similar stability in the number of species remaining in forests after fires. This is in large part a result of the fact that direct mortality due to fire is small (Hakala et al. 1971; Howard et al. 1959; Komarek 1969; Vogl 1973). Bendell also noted that most fires burn unevenly, leaving a mosaic of successional stages in the forest, seldom producing large uniform areas of uniform vegetation. Such mosaic forests provide a variety of environments and wildlife as well as colonists to new burns (Daubenmire 1968; Heinselman 1970; Vogl 1970). While the present studies involved burns of too small a size to test these ideas, future consideration should be given to maintaining the mosaic features of giant sequoia and associated mixed coniferous forests as larger management burns are used.
A number of changes in bird and mammal populations are forecast if fire is reintroduced in the groves on a larger scale, Larger and potentially hotter fires kill more white fir and potentially more sugar pine, the former to heat and the latter to insects. The resultant increase in wood-burrowing insects will result in increased numbers of trunk-feeding birds for several years. This should also result in an increased number of flycatchers and woodpeckers. The magnitude of such changes can be reduced if the burns are kept from becoming much hotter than the experimental burns. The number of Douglas squirrels should not be affected appreciably, while the numbers of California ground squirrels are expected to increase. The latter species is a ground-dwelling form which favors open, well-lighted conditions and which has been noted to increase after logging and burning (Gashweiler 1970; Tevis 1956). The magnitude of the changes in bird and mammal populations will depend on how far back the successional process is pushed by the manipulations, and on the pattern of burning used in treating larger areas of the groves.
Birds, mammals, and giant sequoia reproduction
Birds and mammals exert little effect on giant sequoia seeds either on the ground or on seedlings. The principal reason for this seems to be their small size. A seed weighs but 0.005 gram, and an average of only about 180 fall to each square meter of ground per year. Even then, however, only 5 to 10 seeds fall per day, which is a very low return in food energy for the energy expended gathering, eating and assimilating them. The seed spot tests showed few sequoia seeds were used even when concentrated in groups of fifty seeds. Preference was given by the rodents to the largest seeds in all cases. There is evidence that the animals learned not to spend time on sequoia seeds and to search for larger seeds. Those sequoia seeds eaten were consumed during the first series of tests at a particular site. They were either not uncovered or uncovered but not eaten during the second series at each site, presumably by the animals that visited the site previously. Harvey (Chapter 5) and Stecker (Chapter 7) showed that most seeds on the ground died of desiccation or by insect attack. Douglas squirrels eat some seeds in the cones and cause the shedding of many more.
Few giant sequoia seedlings are eaten by vertebrates. Concentrated masses of these seedlings seldom provide enough biomass to be of interest to an herbivore. There was a higher mortality in boxes of seedlings (25%) than in those seedlings that germinated naturally in the study areas (approximately 1%), presumably because of the differences in their density. Deer occasionally browsed dense groups of seedlings, a phenomenon also noted by Stecker (Chapter 7). Such browsing was haphazard, and the animal usually moved on after a few bites. Some seedlings were crushed by various mammals. Birds do take a few seedlings, again more often when the seedlings grow in dense clusters. This tendency helps explain the views of Fry and White (1930), who suggested that vertebrates have a considerable impact on giant sequoia seedlings after having observed numerous interactions in a nursery setting.
Birds and mammals killed only 1% of the marked seedlings dying from all causes. Their effect was greatest when the seedlings were one to two years old, and especially when they were concentrated in dense groups. Some insects have specialized to support themselves, at least in part, on the small amounts of food energy found in the seeds and young seedlings (Chapters 6 and 7). No bird or mammal is apparently so specialized.
| <<< Previous | <<< Contents >>> | Next >>> |
chap8.htm
Last Updated: 06-Mar-2007