|
NATIONAL PARK SERVICE
Giant Sequoia Ecology: Fire and Reproduction |

|
CHAPTER 9:
DOUGLAS SQUIRRELS AND SEQUOIA REGENERATION
Howard S. Shellhammer
Introduction
John Muir (1911) said of the tree squirrels of the Sierra Nevada:
There are two species here, the large California gray and the Douglas. The latter is the brightest of all squirrels I have ever seen, a hot spark of life, making every tree tingle with his prickly toes. . . a condensed nugget of fresh mountain vigor and valor. . . .
The Douglas squirrel (Tamiasciurus douglasi) (Fig. 59) plays a significant role in the life history and regeneration of the giant sequoia as does no other vertebrate (Chapter 8). Earlier reports emphasized the cone cutting and caching activities of these squirrels (Cahalane 1942; Clarke 1939; Fry and White 1930; Ingles 1965; Murie 1927). Subsequently, Fry and White (1930) and Shellhammer (1966) documented that Douglas squirrels cut large numbers of giant sequoia cones. Fry and White suggested that one squirrel cut and cached "thirty-eight well-filled barley sacks of cones. . . within a period of about twelve days," while Shellhammer (1966) observed one squirrel cut 538 cones from one giant sequoia in 30 minutes. Speculation has prevailed as to the extent that this sort of behavior is common to Douglas squirrels inhabiting sequoia groves, but no detailed studies had been conducted, prior to the present study, to explore this subject.

|
| Fig. 59. A Douglas squirrel, Tamiasciurus douglasi. |
Earlier investigations inferred or believed that Douglas squirrels ate upon giant sequoia cones solely to gain seeds from them, as they do with all other conifer cones (Fry and White 1930; Ingles 1965). The present author questioned this supposition because of the extremely small size of giant sequoia seeds and the green fleshiness of the cones. Hartesveldt and Harvey (1967) reported that it takes an average of about 200,000 seeds to weigh a kilogram (91,200 seeds per pound), each seed (including seed coats and wings) weighing approximately 0.005 gram (Fig. 60). The two hundred seeds in an average cone (Chapter 5) would yield but one gram potential food for the 15 to 30 minutes of effort required to obtain it. Most investigators also assumed that the behavior and biology of Douglas squirrels inside groves of giant sequoias is much like that reported for them elsewhere throughout their range.

|
| Fig. 60. Seeds of the giant sequoia. |
Finally, little is known about the relationship of Douglas squirrels to forest fires. Hatt (1929) described a few of the responses, i.e. escape to tunnels and migration, of pine squirrels (Tamiasciurus hudsonicus) to fires in lodgepole pine forests in the Rocky Mountains of Colorado. There are few other reports, and none in the giant sequoia groves where Hartesveldt and Harvey (1967) and Hartesveldt (1964) have postulated that primeval fires were surface fires which did not often destroy the larger trees. Such fires probably had little impact on the squirrels.
The present study was undertaken to expand our knowledge of the biology of the Douglas squirrel and, specifically, to answer the following questions. What is the impact of the Douglas squirrel on the cycle of regeneration of the giant sequoia? Do Douglas squirrels consume seeds or the cones and, if so, what is their impact? Do Douglas squirrels living in giant sequoia groves behave the same and have similar ecological relationships as do those squirrels who live in non-sequoia coniferous forests? Do moderate-sized manipulations grossly affect the number of squirrels or their biology?
Natural history
The Douglas squirrel is found in the mountainous areas of central and northern California, western Oregon and Washington and southwestern British Columbia. Its close relative, the red squirrel (Tamiasciurus hadsonicus) is found in the New England and Great Lakes states, the Rocky Mountain states, most Canadian provinces, and Alaska. It has been well studied (Hamilton 1939; Hatt 1929, 1943; Klugh 1927; Layne 1952; Muir 1927; C. Smith 1965, 1968, 1970), as has the Douglas squirrel in the northern part of its range (C. Smith 1965, 1968, 1970). The study of C. Smith (1968) provides us with the best account of the typical behavior of both red and Douglas squirrels throughout most of their ranges. Both species are active all year even in the most severe climates despite the fact that they are small and have very high metabolic rates (Irving et al. 1955). They are highly territorial animals which defend their territories throughout the year on an individual basis, except for a very brief period of sexual activity. Most of their territorial defense is by means of vocalizations. These are so efficient as to reduce the amount of their time actually spent defending territories to approximately 2%. C. Smith (1968) speculates that vaginal secretions of the female aide in the coordination of breeding behavior and the associated relaxation of territorial activity on the part of both sexes.
There may be one or two reproductive periods per year, depending on the habitat of the animals. Two to five young are born after a 40 day gestation period. C. Smith (1968) found that young red squirrels stayed in the nest for 50 to 57 days after birth and were not weaned until the 62nd to 64th day. He considered Douglas squirrels to have similar patterns. Young of both species spend two to three weeks before they are completely weaned, exploring the area about their nest upon first leaving it, learning to eat adult foods, and beginning to establish territories. Their mother excludes them from her territory soon after they are weaned, and all individuals in the family group establish defended territories by the time the young are 71 to 74 days old (C. Smith 1968). The major items of food throughout most of the range of either species are the reproductive parts of conifers, deciduous trees and fungi, as well as the cambium of various bushes and pines. They typically build large nests of plant material either in trees or in the ground. The use of such nests and a diurnal pattern of activity allows them to minimize the energy spent on maintaining their body temperature. They cut large numbers of conifer cones and store the cones in caches (large numbers of cones stored in cool, dark or moist places). Such caches are easily defended and can be found and used even under the snow. Indeed, animals eating under the snow are protected from the colder temperatures above the snow level. The sizes of territories in the British Columbia populations studied by C. Smith (1968) in hemlock and in lodgepole pine forests were 0.51 hectares (1.27 acres) and 0.91 hectares (2.24 acres) respectively. The size of the territory and the potential success of its occupant surviving the winter were both related to the amount of food found in it. The function of territorial behavior in these species was described by C. Smith (1968) as ". . . to allow each individual the optimum conditions for harvesting, storing, and defending a seasonal food supply so that it will be available throughout the year. The defense of territories by single individuals allows for a more efficient storage of food than defense of territories by pairs of squirrels."
Methods
Douglas squirrels were observed in North and Trail Areas from 1965 to 1974, Ridge Area from 1966 to 1974. The size of the observation area or plot in North and Trail Areas was 1.8 hectares (4.5 acres) each, and approximately 0.8 hectares (2 acres) for Ridge Area. The observation plots included the manipulated portion of Trail Area and most of the manipulated and control portions of North and Ridge Areas, plus a surrounding buffer zone in each case. The zone of observation was enlarged to 30 hectares (75 acres) in each area in 1974 because of the extremely low and widely dispersed population of squirrels at that time.
Douglas squirrels were censused using the plot census method developed for birds by Kendeigh (1949). Dittoed maps of each area of observation were used, on which to record each day's observation. Eight-power binoculars were used for observations. Field records were kept and correlated with the results of the plot censuses. No attempts were made in the early part of the study to capture and mark animals due to the distances involved in observing them and their wariness of the traps. Observations were made one to two times a week in July and August of 1965, 1966, 1967, and 1974; six days in August and September in 1968; seven days in 1969; four days in 1970, 1971, 1972, and 1973; one day checks in May and October from 1965 to 1968; and one day in October in 1970, 1972, 1973, and 1974 for a total of approximately 650 hours of observation.
Estimation of cone loads on giant sequoias was difficult because the cones remain green and unopened for many years and because each large tree bears a great number of relatively small cones. Cone loads were estimated from the ground for nineteen randomly selected trees in the various areas during 1967 and 1968 using eight-power binoculars. Cone categories used were: 1 = load consisting almost entirely of a few terminal cones; 2 = some concentrations of cones (10-50), few terminal cones or, in some trees, the terminal cones comprising the bulk of the cone load; 3 = few major bundles of cones and then only 20-60 cones per bundle; 4 = numerous large groups of cones (60-200) found on most, or at least a majority of branches with a moderately heavy terminal cone load; and 5 = very heavy load of cones, many large groups of cones with most to all exterior branches filled with cones and with a heavy load of terminal green cones. First year (or new cones) are of a different hue of green than older cones, and their numbers were also estimated on each of the nineteen trees. The cones in one of these trees (the Castro Tree in South Area) were counted from within the tree a year later by Stecker (Chapter 7) and a count for the year in question was calculated by subtracting the numbers of first year cones from that exact count. A cone load factor for that tree was obtained by multiplying the estimated cone load of the tree times its dbh. The number of cones on the same tree was divided by the cone load factor to obtain a multiplier figure which could be multiplied by the cone load factors of other trees to obtain an estimate of the number of cones for each of them.
Early observations during the course of the study revealed that individual giant sequoias were never inhabited by more than one squirrel during any one year. Hence the number of cones cut and chewed per unit time could be estimated by clearing cones from beneath occupied trees in the fall of one year, and picking up and counting the newly cut cones in the next summer. This was done for four trees.
Time spent by cones on the ground was visually estimated by comparing changes in them to the changes that took place in cones previously monitored for known periods of time. Recently cut and chewed cones were yellowish-green, while those down for a few months were rusty brown, and year old cones were grey-brown.
The potential use of sequoia seeds by squirrels and other mammals was checked using the seed spot technique described in Chapter 8.
The potential impact of the squirrels on seeds in cones was checked by shaking out the remaining seeds from 100 chewed and partially dried cones randomly collected in the study areas, and from inspecting 1383 seeds taken from eating places. The viability of these seeds was checked using a snap test.
Potential age-class preference for cones by the squirrels was assessed by comparing 250 green cones removed from six branches of a study tree in South Area with 160 fresh green cones cut by Douglas squirrels from four other trees in the study areas. The age of cones was established by using a dissecting microscope to aid in counting the rings of their peduncles.
The caloric value of giant sequoia seeds and cones was established by burning four samples of seeds and the outer portions of three cones in a standard oxygen bomb calorimeter. Rough estimations of the portions of the cones eaten by squirrels were made by trimming unchewed cones to the approximate configurations of chewed cones. All samples were oven dried to a constant weight at 65-70°C. No additives were needed to insure combustion, as preliminary tests resulted in almost total combustion.
Results
Numbers and densities
The number of Douglas squirrels fully or partially inhabiting the 4.4 hectares in Ridge, North and Trail Areas ranged from a high of 36 animals in the late summer of 1966 to 1 in 1974 (Table 24). The population had decreased to such a low level in 1974 that the area of observation was increased to 90 hectares in which there were eight animals. The aggregate number of territories varied from a high of 11 territories per hectare in Trail Area in 1966 to zero in 1974 (Table 24). The average density likewise ranged from a high of 6.4 squirrels per hectare in 1966 to 0.09 squirrels per hectare in 1974 (Table 24, Fig. 61).
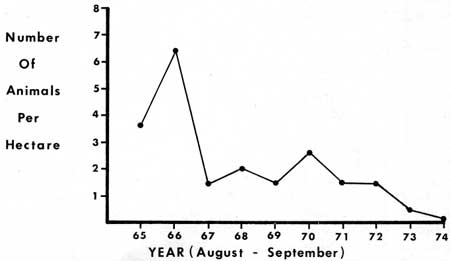
|
| Fig. 61. Mean densities of Douglas squirrels in Ridge. North and Trail Areas (based on August-September data). |
Table 24. Numbers and densities of Douglas squirrels.
| Ridge Area (0.8 ha) |
North Area (1.8 ha) |
Trail Area (1.8 ha) |
All Areas | |||||
| Year | No. animals |
No. territoriesa | No. animals |
No. territories | No. animals |
No. territories | density | |
| per acre | per hectare | |||||||
| 1965 | — | — | 6 | 6.0 | 8 | 7.5 | 1.5 | 3.7 |
| 1966 | 12 | 7.5 | 12 | 10.0 | 12 | 11.0 | 2.6 | 6.4 |
| 1967 | 1 | 1.0 | 1 | 1.0 | 5 | 4.5 | 0.6 | 1.5 |
| 1968 | 3 | 3.0 | 4 | 2.0 | 4 | 3.5 | 0.8 | 2.0 |
| 1969 | 2 | 1.5 | 2 | 1.5 | 4 | 3.5 | 0.6 | 1.5 |
| 1970 | 3 | 2.0 | 7 | 6.5 | 5 | 4.0 | 1.1 | 2.7 |
| 1971 | 2 | 1.0 | 4 | 4.0 | 3 | 2.0 | 0.6 | 1.5 |
| 1972 | 3 | 1.5 | 4 | 3.0 | 3 | 1.8 | 0.6 | 1.5 |
| 1973 | 2 | 0.8 | 2 | 1.0 | 2 | 0.5 | 0.2 | 0.5 |
| 1974 | 0 | 0.0 | 1 | 0.5 | 0 | 0.0 | — | — |
| 1974b | 4 | 4.0 | 2 | 2.0 | 2 | 2.0 | 0.04 | 0.09 |
aTotal of full and partial territories bEach area studied enlarged to 30 hectares (75 acres) in 1974 | ||||||||
The minimum size territory was approximately 0.28 hectares (0.69 acres) and was reached in 1966. This space was the smallest in which both a large giant sequoia and a large white fir could be found, and both appeared to be necessary for a successful territory (Table 25). The numbers of animals dropped to very low levels in 1973 and 1974 during which time territorial defense and stable territories nearly disappeared. Animals wandered over one to two hectare zones with seven to fifteen hectare neutral zones surrounding them.
The average distance between centers of activity varied greatly during the years of the study (Table 25). Average distances in the range of 40 to 60 meters, as in 1966 (Table 25), were associated with intense territorial interactions. Animals with territories centered 150 meters or more apart, or whose territories were much larger than 1 hectare (Table 25), exhibited little territorial behavior towards each other.
Table 25. Average size of territory or area occupied and average distance between centers of activities of Douglas squirrels.
| Ridge Area |
North Area |
Sout Area | |||||
| Year | Territory in hectares |
Distance between centers in meters |
Territory in hectares |
Distance between centers in meters |
Territory in hectares |
Distance between centers in meters |
Mean size territory (hectares) |
| 1965 | — | — | 0.69 | 124 | 0.42 | 75 | 0.52 |
| 1966 | 0.28 | 40 | 0.32 | 61 | 0.28 | 66 | 0.28 |
| 1967 | 0.66 | 102 | 0.68 | 243 | 0.67 | 66 | 0.66 |
| 1968 | 0.48 | 54 | 1.26 | 78 | 1.01 | 65 | 0.82 |
| 1969 | 0.68 | 161 | 0.51 | 192 | 0.66 | 152 | 0.52 |
| 1970 | 0.40 | 151 | 0.64 | 82 | 0.49 | 66 | 0.52 |
| 1971 | 0.68 | 215 | 0.70 | 90 | 1.10 | 140 | 0.76 |
| 1972 | 0.40 | 91 | 1.14 | 80 | 0.83 | 103 | 0.74 |
| 1973a | 9.12 | 200 | 5.47 | 155 | 5.47 | 210 | 6.31 |
| 1974a | 7.50 | 1500 | 15.00 | 1100 | 15.00 | 11.80 | 11.38 |
aTerritories of 1 to 2 hectares were established inside these larger areas. | |||||||
Natural history
No large nests of plant materials, so characteristic throughout most of the squirrel's range, were found in any area of giant sequoias in the ten years of the study. Stecker found a possible nest at an elevation of 82 meters (270 ft) in his study tree in South Area in 1970. It was a hole 20 x 36 cm and contained numerous fecal pellets of Douglas squirrels, some shredded bark and some old pine seeds. It is not known whether this was a summer or winter nest, or both. Numerous growths in old white firs were observed carefully from the ground and none of them appeared to be any kind of nest. The occasional holes in the trunks of giant sequoias, other than the two climbed and examined by Stecker (Chapters 6 and 7), could not be examined due to the difficulty in climbing such trees.
Douglas squirrels apparently used shredded bark to line their nests. Suppressed giant sequoias with their lower trunks shredded are common wherever Douglas squirrels have resided for any length of time. George Lawrence (pers. comm.) suggested that both Douglas squirrels and grey squirrels (Sciurus griesus) shred giant sequoia bark for bedding material. Most of the shredding activity was attributed to Douglas squirrels as most of the affected trees were in or near known territories and because the grey squirrel population was far smaller. The only sighting of animals actually shredding young sequoias was by Karen Shellhammer (pers. comm.), and that was of a Douglas squirrel shredding a small tree among a group of previously shredded trees. Trees used for shredding were always small (10-20 m tall), suppressed and growing in dense groups. A few to most of the trees in such a group were shredded over 50 to 80% of their diameters and from 10 cm to 8 m off the ground. They were stripped of their bark but not their cambium and were still living. Occasionally one of the smallest trees had enough cambium removed so as to kill it.
Precise data concerning reproductive rates and longevity were difficult to collect as the animals were not marked. It was possible, however, to gain a general idea of these patterns because many of the animals maintained stable territories for several years, especially in years of moderate to high densities. The mean life span of these animals was estimated to be approximately two years, although this figure is influenced greatly by the high mortality in 1966-67 when 30 of 36 animals apparently perished.
Second litters were seen outside the study areas in 1965, 1966, 1969, and 1972, and in summers following mild winters and early growing seasons. Douglas squirrels are not weaned until 102 to 104 days following conception (C. Smith 1968) and usually established territories in late July and August. Second breedings apparently did not occur in the study areas in any year and were infrequent in the Redwood Mountain area. No accurate measurement is possible for the average number of young produced per year.
Foods
The major item in the diet of Douglas squirrels in the study areas was the cones of giant sequoias. They were taken nearly year round in years when the squirrels were dense and primarily in the fall, winter and late summer in other years. The squirrels switched to white fir, sugar pine and/or ponderosa pine cones as they matured in the last part of August or early September each year. The cone type consumed depended upon the relative abundance of its cones during the fall. White fir cones were extremely abundant and used heavily in 1973 but were absent altogether in 1974. The few squirrels that were present in 1974 turned to ponderosa pine, as the sugar pine crop was also poor and patchy in distribution. The squirrels usually spent two weeks feeding on these alternative conifer cones but never were seen to cache them. Douglas squirrels cached only giant sequoia cones in the study areas, whereas they cached the cones of other species of conifers in nearby nonsequoian forests. This phenomenonrep resents a primary departure from Douglas squirrel behavior in the rest of their range.
Douglas squirrels harvested hazelnut (Corylus rostrata) and mountain alder fruits (Alnus tenuifolia), the latter to an undetermined degree, each August and September. Fungi of a variety of species were eaten throughout the summer and early fall and some were stored for winter use.
Those Douglas squirrels observed eating giant sequoia cones took about 15 to 30 minutes to chew away the outer portion of the cone. They were never observed to eat brown, dried cones or those with several to many brown scales on them. Squirrels were observed eating cones while at the base of a tree or on branches 3 to 10 m above the ground. Those eating cones in the upper branches of 85-100 m tall giant sequoias were unobservable. Cones were picked up with both forefeet, with one end held higher than the other. The cone was rotated with the forepaws as the outer portion of each green scale was chewed off and dropped. The tips of several scales were trimmed before the squirrel concentrated on individual scales. The upper and lower surfaces of each scale were scraped repeatedly before the animal switched to the next scale. Some of the seeds attached at the base of the scale were consumed in this process, but most of the six to eight seeds attached to each scale remained attached and undamaged or were spilled from the cone in a relatively undamaged condition along with some of the scrapings from the scale. The pattern of rotate, trim and chew was continued until most of the scales of the cone had been worked, and then the cone was dropped. Some squirrels had "signatures" in relation to the amount of green cone they left uneaten (i.e. one ring of scales, two, three, etc.) (Fig. 62). A small percentage of squirrels stripped the cone almost to the main axis leaving an object resembling the core of a small, well-eaten apple. Squirrels groomed their faces with their hindfeet and cleaned their whiskers by rubbing their faces on a branch of a tree.

|
| Fig. 62. Sequoia cones: dry, brown cone on left; green, fleshy cone in center; cone chewed by Douglas squirrel on right. |
Chewed and partially dried cones picked up from the ground were found to have a small percentage of their seeds, and most of these were relatively undamaged physically. Two hundred and nine seeds (67%) of 310 shaken from 100 dried cones were whole and undamaged. Thirty-eight percent (525 of 1383) of the seeds collected from an eating place, or "kitchen midden," were whole, whereas 62% were physically damaged, although most of the damage was to the sterile wings of the seeds. The viability of such seeds was reasonably high (29%) using a snap test. This level of viability compares favorably with the results of controlled drying tests of seeds and indicates that the saliva of the squirrels does not appear to damage the seeds.
Cone cutting behavior
Douglas squirrels used three different patterns of cutting giant sequoia cones. They cut large numbers of cones in a short time, small numbers intermittently over a long time, or they cut individual cones and ate them. The most dramatic of these was the cutting of large numbers of cones in brief periods of time. Shellhammer's report (1966) of one squirrel's cutting 538 cones from one tree in 30 minutes is the most stunning example of this pattern. The squirrels cut rapidly and yet selected for certain age classes of green cones and excluded brown and nearly all partially brown cones. Cones cut in such periods were cached in several ways. Some animals buried 2 to 5 cones in little pits dug in the ground within ten meters of the base of a giant sequoia or about its base where the duff was especially thick, Other animals cached cones in the dark, cool, deeper parts of fire scars. The fire scar of one giant sequoia contained 538 cones in one scar and 189 in another. The caching of 500 to 750 cones, regardless of the type of cache, usually required three to four days. A third type of cache was found in damp or moist areas. One such cache in Trail Area in 1966 contained an estimated 725 cones in three groups along a seasonally moist area. There are numerous reports of similar types of caches of other conifer cones in nonsequoian forests. Shaw (1936) reported on the caching activities of various subspecies of Douglas squirrels and noted that such cool, moist places were extremely effective in keeping cones green and moist. Douglas squirrels, however, do not cache their cones in creeks. Cones end up in water courses by accident when squirrels cut trees near them. Cones are cut from the upper regions of giant sequoia trees and fall in 180° to 360° arcs about their bases. The squirrels subsequently place those cones that have fallen onto the forest floor in caches. They do not attempt to retrieve cones from creeks, and the currents tend to deposit cones in low spots, hence the appearance of "aquatic caches."
A second pattern of cone cutting occurred at much lower levels of intensity. Cones were cut in small numbers per period of cutting over longer periods of time, and each group was eaten or cached before another group was cut. Cones were cached poorly or, often, not cached at all when the numbers of squirrels were low. New cones were cut when the last few cones of the group cut previously began to dry out and were no longer edible.
The third pattern was one in which squirrels cut, ate and dropped individual cones from the tree itself. This pattern was quite variable, being exhibited by many young squirrels and by many adults during fall and winter. The numbers of adults exhibiting the last two low intensity patterns increased in years when the populations of squirrels were low.
The patterns of cone cutting and territorial behavior will be discussed as they appeared in years of high, moderate to low and very low numbers of squirrels.
Behavior in years of high densities
The summer of 1966 produced the highest densities of Douglas squirrels observed during the length of the study (Table 24, Fig. 61). Territorial activity was intense, with the highest frequency of vocalization per animal demonstrated. Many chases took place, and several of the chases observed in August ended in physical contact and cries of pain on the part of the bitten animals. Alarm calls directed towards the investigators were easily elicited and at greater distances than in any other year.
Squirrels were nearly equidistant from one another, with a giant sequoia as the center of each territory. Each territory contained one or more large, old white firs which often served as access to the big tree. Squirrels restricted their cone-cutting activities to a single sequoia, even in territories where two or more sequoias were present. Cones were cut in great numbers. In addition to the bout described by Shellhammer (1966), individual squirrels were observed to cut 650 cones in 55 minutes, 206 in 17 minutes, 290 in 3 hours, and 997 in 5-1/2 hours. Another animal cut 1,430 cones in three bursts of several hours spread over a three-week period. Almost all cones cut were cached about the bases of giant sequoias or in wet areas. Most of the cones cut were eaten on the ground, at kitchen middens, or on the lower branches of trees.
Douglas squirrels were intolerant of other species of rodents, especially chipmunks (Eutamias spp.). Chipmunks were chased from caches of cones, while California ground squirrels (Otospermophilus beecheyii) were threatened but not attacked by the Douglas squirrels.
Behavior in years of moderate to low densities
The summer of 1968 followed the second driest and mildest winter of the period of the study. The numbers and densities of Douglas squirrels were approximately one-third that of 1966 (Table 24, Fig. 61), and the size of their territories was approximately three times greater (Table 25). Territories included five or more giant sequoias, and two or more of them were cut by most squirrels during the year. All types of calls were low in number and most animals gave fewer territorial calls than in 1966. Juveniles had much larger unoccupied and undefended spaces between existing adult territories into which they could move to set up their own territories. Adults actively defended a central core of 0.7 to 1.0 hectares about the tree or trees in which they were residing and weakly defended a narrow buffer zone around the core.
Few squirrels cached large numbers of cones. One squirrel cached 350 cones in an old, downed sequoia, but this animal exhibited similar behavior each year, unlike most other squirrels. Several animals cut a majority of first year cones, although this was the only year of the study that it occurred to any extent. Animals cutting 30 cones or more at one time seldom cut any first year cones. No long bouts of cone cutting were observed, nor were any large caches of cones found.
Sugar pine and white fir cones were used heavily in late August and September, as they were in 1966.
Behavior in years of very low densities
The populations of Douglas squirrels were about 70 times lower in the summer of 1974 than they were in the peak year of 1966 (Table 24, Fig. 61). The population had dropped to the second lowest level during the period of the study in 1973 following a very severe winter. The winter of 1973-1974 was average for the period of the study but was followed by an even further drop in the numbers of squirrels. Only one animal was found in any of the study areas in the summer of 1974, hence the decision was made to enlarge each area of observation to 30 hectares (75 acres). Eight animals resided in the 90 hectares comprising the new study areas.
Calling nearly disappeared throughout the summer, with the exception of a few territorial calls in August. Animals did not give alarm calls to the presence of investigators unless they were surprised at close distances. Only one of the eight animals cut cones in any numbers, cutting an average of 200 cones a day in late August. It was also the most vocal animal.
Territories were essentially nonexistent as the center of activities were 1100 to 1500 meters apart (Table 25). No single giant sequoia was used for more than several weeks to a month with the exception of the animal described previously, and that animal became established only after residing in three other sequoias for brief periods of time. The average squirrel used seven trees from the winter of 1973 to late August in 1974. Most cones were cut and eaten singularly in the upper parts of the trees and then dropped. Each of the squirrels, however, cut 150 or more cones a day on three or four days in July or August. These were not effectively cached and in several cases not even eaten. Some cones were superficially cached on the top and along the sides of downed trees, and no large caches were found, with one exception. Almost all of the cones in these occasional bouts of cutting dried out on the ground and became inedible. The general pattern was for an animal to eat cones in a tree for several weeks, cut a number of cones, eat a few of them and then move on to another giant sequoia 20 to 200 meters away.
The white fir cone crop failed and there were few sugar pine cones. Occasional sugar pine cones were eaten early in the summer, an unusual activity when compared to other years.
Age class preference for cones by squirrels
Most (84.9%) of the cones cut by Douglas squirrels were in the 2 to 5 year age classes, while only 46.3% of the cones taken from the study tree were of those age classes (Table 26). The squirrels took very few older cones, as was the case with almost every group of cut cones examined throughout the study.
Impact of Douglas squirrels on cone loads
The giant sequoia used as a reference point (Castro Tree in South Area) was estimated to contain approximately 18,000 cones in 1969 based on a count made from within the tree by Stecker and Shellhammer. A more accurate figure of 18,411 (for 1969 cone crop) was obtained in 1970 by subtracting the number of first year cones (20,697) from a cone by cone count of the tree (39,508) done by Stecker in that year (Chapter 7). The estimated number of cones in the Castro Tree (18,411) was divided by its cone load factor (44) to yield a multiplier figure (418.43). This figure was multiplied by the cone load factor of each of the other trees in the sample to obtain a rough estimate of the numbers of cones in each of them. Using this method the average number of cones on 19 trees was 14,117 (5,021 to 25,106). The numbers of cones on large giant sequoias on Redwood Mountain varied from a few hundred to an estimated 55,000 or more. Stecker's (Chapter 7) as well as my own estimates indicate that 60 to 66% of the cones on an average tree are green and 33-40% are brown at any particular time, except after such exceptional years like that in 1969-70 when great numbers of first year cones were produced. Approximately 16% of the green cones are first year cones (Table 26), although a much higher percentage can be present following favorable wet years (Chapter 7). Using these figures, the number of green cones on an average giant sequoia in the study areas was estimated to be approximately 9,000, of which 1,500 are added each year, and a similar number was expected to turn brown or be knocked off by snow or wind.
Table 26. Age class distribution of giant sequoia cones versus age class preference by Douglas squirrels.a
| Age class of cones in years |
Percentage of 160 cones cut by squirrels |
Percentage of 250 cones from Tree 15, South Area |
| 1 | 3.8 | 16.0 |
| 2 | 23.1 | 5.2 |
| 3 | 38.8 | 19.5 |
| 4 | 15.0 | 15.6 |
| 5 | 10.0 | 10.0 |
| 6 | 5.0 | 5.0 |
| 7 | 3.1 | 10.0 |
| 8 | — | 5.2 |
| 9 | — | 2.4 |
| 10 | 0.6 | 4.8 |
| over 10 | 0.6 | 7.2 |
aA chi-square test showed the differences to be highly significant (p.<.001); x2 = 116.3, d.f = 10. | ||
To get an idea of the magnitude of the number of cones produced in the four study areas during the ten years of the study, one can consider the cone loads estimated in 1967 and 1968 as a starting point, and that 16% of that green cone load would be added per year for a very conservative figure. The estimated starting load of green cones on the 65 largest giant sequoias in and near Ridge, North and Trail Areas was 578,110, plus 92,495 added for each of nine years, for an estimated ten year total of 924,950 cones. This works out to about 9,000 cones/ha/yr being produced. If a steady state of cone production existed, seed production would be about 1,800,000/ha/year.
Most (86.6%) of the 65 largest giant sequoias in the study areas had cones cut from them by Douglas squirrels during the ten years of the study (Table 27). Cones from two trees were cut in nine of the ten years, but relatively lightly in the last three of those years. Eighteen trees were cone-cut heavily (2,000 cones/year or more) from one to six years (Table 27). Each tree was estimated as being lightly cut (1 to 100 cones, average of 50), moderately cut (100 to 1,000, average 500) or heavily cut (1,000 to 3,000+, average of 2,000). An estimated 135,505 cones were cut from the 65 trees in the ten years, or 2,085 cones per tree in ten years. This is an average of 209 cones per tree per year, but this figure rises to 251 cones per tree per year if uncut trees are excluded.
Table 27. Giant sequoias in which Douglas squirrels cut cones.
| Trees cut at any level of activitya |
Trees cut heavily; 2,000 cones or more | |||
| Number of years | Number of trees | Percentage | Number of trees | Percentage |
| 0 | 10 | 15.38 | 47 | 70.76 |
| 1 | 18 | 27.68 | 6 | 9.32 |
| 2 | 10 | 15.39 | 2 | 3.08 |
| 3 | 10 | 15.39 | 4 | 7.69 |
| 4 | 5 | 7.69 | 3 | 4.62 |
| 5 | 2 | 3.08 | 2 | 3.08 |
| 6 | 2 | 3.08 | 1 | 1.54 |
| 7 | 4 | 6.15 | — | — |
| 8 | 2 | 3.08 | — | — |
| 9 | 2 | 3.08 | — | — |
| Totals | 65 | 100.00% | 65 | 100.00% |
aFrom 50 cones or more per year to more than 2,000 cones per year. | ||||
Coefficients of correlation between cone load (as of 1967-68) and the number of cones cut in the ten year period for Ridge, North and Trail Areas were 0.40, -0.36 and -0.25 respectively. Hence there was no obvious tendency by squirrels to select trees with high cone loads. Only six trees had 10,000 or more cones cut from them during the period of the study. Two of these trees had light cone loads (5,000 and 7,000), two were in the third heaviest category (of five) and two were in the second heaviest category. One tree was used for seven years in a row; another for six years. Most trees that were used heavily were cut in two series of years with several years of disuse in between. A somewhat random alternation of tree use was produced by a combination of: young squirrels establishing territories in the free spaces between existing adult territories; the creation of free spaces between the young squirrels' territories by the subsequent deaths of the adults; fluctuations in the size of territories; and the number of trees used within each territory. Few trees were used repeatedly for long periods of time because of these factors.
Caloric studies of cones and seeds
The mean caloric value of giant sequoia seeds as measured by bomb calorimetry was 4,738 cal per gram d.w. and that of the outer portions of the cones was 4,690 cal gram d.w.. The outer portions of four cones (mean weight = 59.3 g) were trimmed to approximate chewed cones. The trimmings averaged 33% (19.7 g) of the wet weight of the cone and were dried to an average of 10.2 grams. Using C. Smith's calculations (1968) as a model, 9.5 grams of what was lost were considered to be water or plant juices and 1.2 grams of the dried trimmings were estimated to be indigestible supportive materials, leaving 9.0 grams, or 15% of the total wet weight, of each cone as dry weight material available to a squirrel.
Net energy available to a squirrel also was estimated using the guidelines of C. Smith (1968) for both Douglas and red squirrels. He suggested they digest a high percentage of their food and that the calorigenic effect of assimilating it was low. He used a figure of 75% of the ingested food energy as an estimate of net energy, while 70% is used in this study, as the author estimates the squirrels are ingesting more crude fiber. Measurements of 65 cones from various areas yielded an average weight of 46.2 grams. The maximum energy gained from eating such a cone is calculated as 46.2 g x 0.15 (% dry wt [d. w.] of chewed materials) x 0.7 (% of ingested material assimilated) x 4,690 cal per gram d.w. = 22.8 kg calories per cone. This figure is a liberal one; most animals likely eat less of the cone than the estimated trimmings as they discard the tips of the scales and spill some other tissues. A squirrel eating such a cone in 20 to 30 minutes could obtain 1.14 to 0.76 kg cal per minute, whereas the energy obtainable from the average of 200 seeds for such a cone is far lower. Two hundred seeds in an average cone yield 1.4 kg cal, or from 0.07 to 0.05 kg cal per minute over 20 to 30 minutes. Hence a Douglas squirrel might gain 10% more food energy and a possibly more nutritious food by eating all the seeds of a green sequoia cone in addition to the outer portions of the cone, but very little energy (0.05 to 0.07 kg cal per minute) if it tore a cone apart solely to obtain the seeds. The latter intent would provide about one-fifteenth the energy gained by eating the scales, assuming that both or either activity took an average of twenty minutes.
Douglas squirrels cut 2,719 and 3,750 cones from two different giant sequoias in 318 days during 1968 and 1969 for an average of 8.6 and 11.8 cones per day. These figures are a bit misleading as it is probable that fewer cones were eaten in some months when other conifer cones and fungi were available. More cones were eaten most likely in late winter and early spring when all other food sources were used up. Squirrels also removed 2,082 and 1,583 cones from other marked trees during 1971 and 1972, but the number of days could not be determined. It is estimated, using these figures, that a squirrel might cut and eat on 2,500 to 4,500 giant sequoia cones per year in addition to much smaller numbers of white fir or sugar pine cones plus fungi and assorted other foods.
Weather factors
Measurements of temperature and snowfall taken from Grant Grove approximately 300 meters higher and 5.5 kilometers away from the field areas showed a winter average maxima 5°C colder and average minima 3°C higher than in the study areas, while summer average maxima and minima were the same. Two measures each of heating degree days and snowfall are shown on Table 28. Heating degree day measurements are standard U.S. Weather Bureau figures for a number of degrees (in °F) below 68°F each day on a monthly basis. Sixty-eight degrees F is also the critical temperature below which the Douglas squirrel has to expend in excess of basal metabolic requirements to maintain a constant body temperature (Irving et al. 1955). Irving et al. (1955) determined the figure for animals belonging to a subspecies of red squirrel in Alaska, but both C. Smith (1968) and the present investigator consider it to be similar in Douglas squirrels. The relationship between the average density of the squirrels in Ridge, North and Trail Area (from Table 24) and number of heating degree days both per year and in the colder period from January through June each year can be seen on Table 28. The coefficients of correlation in comparing squirrel densities and the two measures of temperature were -0.53 and -0.59 respectively. The correlation coefficient between total snowfall per year and density was much lower, at -0.34, although there was a stronger relationship between the lateness of the snow and a drop in the density of the squirrels. The numbers of squirrels decreased from the level of the preceding summer in 1967, 1969, 1971, and 1973 when the snows were heavy and late. The snow was very late on the ground in the spring of 1967 when 363 cm of 681 cm total snowfall (143 of 268 in) fell in April. The winter of 1970-71 was average for the period of study, but the snow was distributed more or less equally when 29 cm of 518 cm (11.6 of 203.6 in) fell as late as May. Decreases in numbers of squirrels therefore appeared to be correlated with a variable combination of winter temperature, total snowfall and the lateness of the snow pack (and associated lower than usual temperatures).
Table 28. A summary of weather factors and changes in densities of Douglas squirrels.
| Year | Avg. density of squirrels per hectarea |
Compared to the previous summer |
Heating degree days per yearb |
Heating degree days, Jan.-June |
Compared to previous year |
Total snow per year cm (in) |
Month(s) of heaviest snowfall |
| 1964-1965 | 3.7 | 7104 | 4482 | 602(237) | April | ||
| 1965-1966 | 6.4 | more | 6601 | 3955 | warmer | 559(142) | December |
| 1966-1967 | 1.5 | less | 7021 | 4598 | colder | 681(268) | April |
| 1967-1968 | 2.0 | more | 6390 | 4074 | warmer | 292(115) | December |
| 1968-1969 | 1.5 | less | 7237 | 4520 | colder | 856(337) | February |
| 1969-1970 | 2.7 | more | 6619 | 4231 | warmer | 227(107) | March |
| 1970-1971 | 1.5 | less | 6874 | 4381 | colder | 518(518) | December |
| 1971-1972 | 1.5 | no change | 6741 | 3805 | warmer | 391(145) | December |
| 1972-1973 | 0.5 | less | 7537 | 4469 | colder | 765(301) | Oct.-Feb. |
| 1973-1974 | 0.09 | less | 6827 | 4194 | warmer | 498(196) | Jan.-March |
| Average | 6895 | 4271 | 521(205) | ||||
aFrom Table 24 for the summer of the second year listed. bWeather data from Grant Grove, Kings Canyon National Park. Heating degree days = number of degrees (in °F) below 68°F each day during a particular period of time. | |||||||
Responses of squirrels to manipulations
Planned surface fires, litter removal and pile burning on plots of 1 to 2 hectares did not appear to have any effect on the numbers of Douglas squirrels. No major differences in numbers were seen between the treated and control areas in Ridge, North or Trail Area following treatment. Douglas squirrels apparently were unaffected by larger prescribed burns carried out on Redwood Mountain in 1969. Douglas squirrels were observed to move across the manipulated area soon after the ashes cooled, and cone cutting commenced several days after the fires. The survival of these animals over a longer period of time was not studied to the extent of those in our study areas, hence the long term effects of large scale manipulations could not be assessed.
Discussion
The most obvious difference between Douglas squirrels living in giant sequoia forests and those inhabiting other types of forest is that they store giant sequoia cones to the exclusion of all other species of conifers. They eat the seeds of other conifers when they are available but they do not cache any of them. The range of foods eaten, other than conifers, is otherwise quite similar in scope to that reported by C. Smith (1968) in British Columbia, although sequoian squirrels do not appear to store hazelnuts or fruits of mountain alders.
No large tree nests were observed in a giant sequoia forest during the period of the study. Douglas and red squirrels are known to nest in hollow trees, hollow logs, outside tree nests of grass or foliose lichens, or holes in the ground (C. Smith 1968). Their nesting material may include grass, shredded bark or lichens; circumstantial evidence indicated that sequoian squirrels used considerable amounts of shredded sequoia bark. No large ground nests were discovered, and the one tree nest discovered by Stecker (pers. comm.) in the upper part of a giant sequoia offered its occupant little protection from weather and predators.
Large numbers of sequoia cones were stored and eaten at caches in years of high squirrel densities. Most cones were eaten in the trees in winters when the numbers of squirrels were low. This must have exposed the animals to lower winter temperatures for longer periods of time than C. Smith (1968) suggested was the case in British Columbia. He suggested they spent much of their time in their tree nests or beneath the snow eating cones stored there in large caches. Michael Smith (1968) reported that the Alaskan red squirrels also spent much of their time beneath the snow, eating and resting during the coldest periods of the winter.
There was no major movement of squirrels from one area to another. Such movements have been reported for Douglas squirrels from other regions, where they moved from areas of low to those areas of high food availability (Merriam 1884; Klugh 1927; McKeever 1961; C. Smith 1968).
Sequoian squirrels generally had one litter per year with a few exceptions tn the drier, warmer years. C. Smith (1968) noted that Douglas squirrels in the Pacific Northwest west of the Cascades generally had two breeding seasons. There the group of young produced by later winter matings were fed on cached cones, while those of late spring matings were fed on the new season's cones.
Sequoian Douglas squirrels did act quite differently than those from nonsequoian forests. No animals were studied along the edge between a dense grove and a more diverse Transition or Canadian Zone forest to see if their preoccupation with giant sequoia cones remained. This species is quite adaptive behaviorally, and many of the patterns observed in this study must have been shaped by imprinting and other early learning of the young squirrels.
Territoriality
The territorial behavior of sequoian Douglas squirrels is different than that of squirrels living in other forests, especially in more northerly ones. The territories reported by C. Smith (1968) were centered about a group of cone-producing trees and one to several caches. Kemp and Keith (1970) reported that the closely related red squirrel in New York state had two types of territories. One was a prime territory, which was similar to that described by C. Smith (1968) in that it contained a group of conifers producing a good supply of seed and was defended all year. The second type of territory was the winter food cache, which was defended only in the winter. These territories are all centered about a seasonal food supply. C. Smith (1968) defined the function of territorial behavior as allowing "each individual the optimum conditions for harvesting, storing, and defending a seasonal food supply so that it will be available throughout the year." Brown (1964) suggested that some resources are distributed in time and space so as to be economically defensible, and conifer cones certainly are one such resource. The dispersion of most Douglas and red squirrels appears to be a combination of a reasonably uniform distribution of food and of the agonistic behavior of the squirrels themselves. Agonistic behavior is described here as a collective term for attack, threatening, retreat, and/or submissive behaviors. Douglas squirrels usually have low enough densities that their highly aggressive behavior causes them to be dispersed in a more or less regular pattern. High densities may force some squirrels into less suitable habitat (Brown and Orians 1970), and in nonsequoian forests this may end in the death of the individual (C. Smith 1968). Agonistic behavior continues to play a very important role in giant sequoia forests even though the food supply is, for all intents, a constant one.
The animals in the present study were quite regularly spaced at all population densities except the very lowest levels. Such a regular pattern of dispersion is quite important as it tends to minimize the number of possible competitors about any particular animal. This type of territorial defense is optimal where the numbers of individuals are moderately low and a food resource is stored in the center core of a territory (Hamilton and Watt 1970). Such territories become less efficient with two animals in them as the maximum radius will have to be doubled to provide food for the two animals. The total locomotive effort required to obtain all the resources in such a territory will increase by the same amount (C. Smith 1968). Individual territories are most efficiently maintained if their perimeters do not have to be patrolled by the occupant. Douglas squirrels use vocalizations directed from the core of their territories for most of their territorial defense. C. Smith (1968) noted that 76% of their defense was vocal, and only 24% involved chasing intruders. The squirrels he studied were so efficient as to spend but 1 to 2% of their time defending territories.
The territories in giant sequoia forests served as something more than a food source to Douglas squirrels in years when their numbers were high. A giant sequoia, which always had more than one year's supply of cones on it, became the focal point of an animal's agonistic behavior in keeping other squirrels a certain distance away from it. The density never reached the point where each large sequoia was inhabited by more than one squirrel, and it is questionable whether such a situation could occur. The large sequoia approaches being a supernormal stimulus for aggressive behavior in these animals, as well as the center of a territory.
The behavior of these squirrels at low densities is equally intriguing. Territorial behavior essentially disappears when animals are separated by more than 200 to 300 meters. The use of one sequoia becomes infrequent and animals move between sequoia trees. They cut most of their cones in the trees and must increase their exposure to lower winter temperatures, in contrast to the situation at high densities where they apparently eat at caches. Such behaviors would appear to decrease the chances of survival of an animal in years of low population densities.
Impact of Douglas squirrels on giant sequoias
Douglas squirrels have a moderate to slight impact on giant sequoias. Most trees in a hectare are cone-cut in any ten-year period but few trees are cut heavily. An occasional tree with a light cone load may become the core tree of a long-lived female and may have most of its cones removed from it for five to six years, but such a situation is rare. Such trees are seldom used by two generations of squirrels and hence have a good chance of producing new loads of cones, and in a few years will be up to average for cone load.
The cutting of giant sequoia cones is nearly a year-round activity and as such may aid in the regeneration of the grove. Most of the seeds dropped either from the trees or on the ground are relatively undamaged, although those seeds dropped during the summer almost certainly die because of desiccation (Chapter 5) or arthropod damage (Chapter 7). Those seeds dropped just before the first snow or just as the snow melts may contribute to sequoia regeneration, assuming other conditions are near optimum. The rate of success of sequoia regeneration due to the activities of Douglas squirrels must be extremely low, but it is partially balanced by the very slow rate of replacement of the tree.
Impact of manipulations on Douglas squirrels
The numbers and behavior of Douglas squirrels do not appear to be affected by treatments of the type and size described in this study. Little inference can be made as to the potential effects on them of large scale managed fires.
Sequoia cones, weather, and the survival of Douglas squirrels
Douglas squirrels are extremely active animals, both behaviorally and metabolically. Applying the figure of Irving et al. (1955) for the Alaskan red squirrel to Douglas squirrels, they have 1.76 times the basal metabolic rate expected from the relationship between weight and metabolic rate for most animals. If this rate was applied to Douglas squirrels, a male would have enough energy to perform heavy labor 24 hours a day or to put on several grams of weight a day. A female could work as hard and still supply adequate food for her young. C. Smith found that adult squirrels of 243 to 289 grams ingested between 109.7 and 322.3 kcal/day during the summer months. He calculated the daily energy requirements for the colder half of the year to be two-thirds that of the summer period because squirrels in his area readily ate stored materials from large caches and spent most of the rest of the time in their nests. The squirrels in sequoia groves are more likely to expose themselves to winter temperatures because of the behavior patterns previously discussed. They must survive the hardest and coldest parts of the winter and early spring eating on the cones of giant sequoias, as they do not store other conifer cones and have most likely used up any stored fungi by that time. A 250 g male could get 120 kcal/day eating the outer portions of 5 to 8 cones, if eaten under ideal conditions. The number of cones required depends on their size, the temperature, where they were consumed, and the insulation of the animal's nest. Just manipulating the cone while eating it is moderately hard work for the squirrel and requires a considerable output of energy. Eating such a food source in cold temperatures may greatly reduce the net gain of energy involved in the process. Such a constant but just adequate food supply results in an inverse relationship between survival and the severity of winter weather. Cold winters, late snow packs and late springs were correlated somewhat strongly with decreases in numbers.
No estimate can be given for the level of predation by mammalian predators during the winters. Pine martins (Martes americana) are considered by Ingles (1965) to be predators on Douglas squirrels and may have contributed to mortality, especially in the winter of 1966-67.
There must be other factors in addition to those already discussed. These animals, for example, may undergo cyclic fluctuations, although the time frame of the study was too short to show if this was the case. It is difficult to explain the drop in numbers to near zero in 1974 from the very low population in 1973 on the basis of weather alone. The winter of 1973-74 was the most typical one of the ten-year period, and much milder than the harsher winters that preceded other declines. The decrease in cone cutting behavior, however, that was associated with low densities may have made the animals more prone to winter death.
In summary, the Douglas squirrels living in groves of giant sequoias differ considerably from their counterparts in other types of forests. They do not build large tree nests nor store the cones of conifers other than those of the giant sequoia. Cones are cut in greater numbers in years of high densities of squirrels, and the cones become almost more of a territorial object than a food source. The animals eat the fleshy outer parts of the green cones and some, but not significant amounts, of the tiny seeds. They may cut as many as 10,000 cones or more in a five-year period from a single tree, but their overall effect is slight in that they cut but 200 cones on the average per tree per year. This number is far less than the estimated 1,500 new cones added to the average tree each year. Many to most of the seeds in the cones are released relatively unharmed as the cones are eaten, and may contribute in a minor way to regeneration during brief periods of the year. The numbers of Douglas squirrels living in a grove fluctuate, apparently in response to weather and other unexplained factors. Giant sequoia cones appear to constitute a near minimal diet in most winters. Squirrel numbers do not seem to be affected by the use of fire and associated manipulations to the extent used in this study.
| <<< Previous | <<< Contents >>> | Next >>> |
chap9.htm
Last Updated: 06-Mar-2007