|
NATIONAL PARK SERVICE
Mountain Goats in Olympic National Park: Biology and Management of an Introduced Species |

|
APPENDIX A2:
Subalpine and Alpine Plant Communities
E. G. Schreiner
During 1981-90, we sampled subalpine and alpine vegetation to describe and classify plant communities (Appendix A1). Our study represented vegetation on mountain goat summer range, except that we did not sample the Mount Olympus massif (mostly rock, snow, and ice) or the central Bailey Range. Sixteen plant communities were identified at elevations above 1,520 m (see text Fig. 9; Table A1).
Table A1. Mean cover (C) and percent frequency (F) of dominant
species in subalpine and alpine communities.
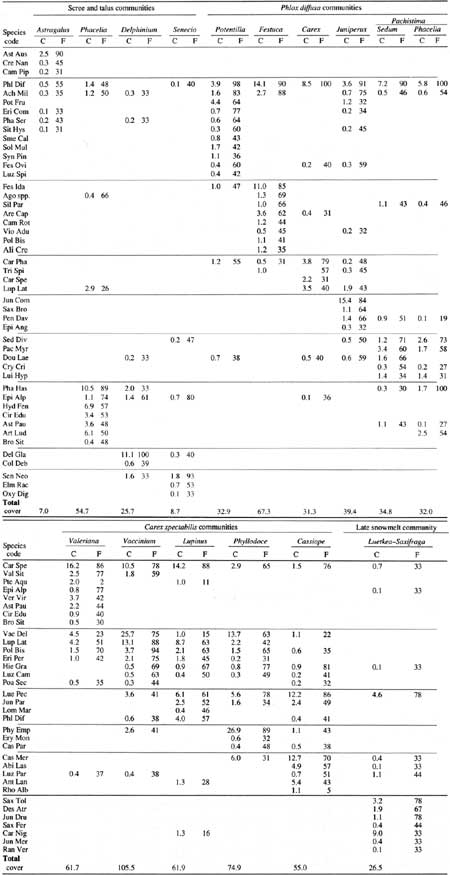
(click on image for a PDF version)
Scree and Talus Communities
Scree and talus slopes are often encountered in the Olympic Mountains because of the steep, rocky terrain and predominance of soft, sedimentary bedrock. Four plant communities were associated with scree and talus. One occurred on steep, bare slopes near ridges; the others were found mainly on colluvial debris fans below massive rock outcrops (i.e., mountains). Distinguishing features of the scree and talus communities were high proportions of rock fragments in the >2-mm—4-cm and 5—10-cm classes; steep (30-35°) slopes; unstable substrates; and 57-97% mineral soil and rock cover (Table A2). Aspect and elevation further distinguished these four communities.
Table A2. Mean environmental and substrate characteristics by
community type. Substrate data represent mean percent cover.
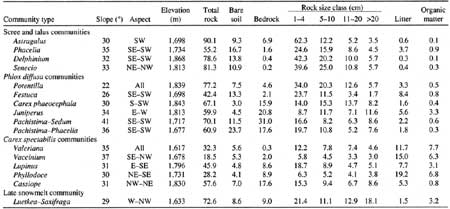
(click on image for a PDF version)
The driest of the four scree and talus communities was dominated by the endemic Astragalus australis var. olympicus. Present only in the driest region of the park, this community occurred on soils with pH 6.0 or higher (Kaye 1989). Total plant cover was 7%.
The three colluvial fan communities were differentiated by aspect, elevation, and substrate. Slopes dominated by Phacelia hastata occurred on dry south exposures averaging 1,737 m elevation; those dominated by the endemic Senecio neowebsteri occurred on slightly higher (1,814 m) and cooler northwest to northeast exposures (Table A2). Sites dominated by Delphinium glareosum were found on south- to northwest-facing exposures at an average elevation of 1,860 m. Most rocks at the surface in the Senecio and Delphinium communities were in the >2-mm—4-cm and 5—10-cm size classes; a higher proportion of the substrate was bare mineral soil in sites where Phacelia dominated. Total plant cover was 9% in Senecio, 25% in Delphinium, and 55% in Phacelia communities.
Phlox diffusa Communities
Six plant communities contained Phlox diffusa as a dominant or codominant. These are differentiated on the basis of slope and substrate characteristics. Slope ranged from 22° in the Phlox—Potentilla community to 41° in the Phlox-Pachistima type (Table A2). Two communities with relatively low slope, (Phlox—Potentilla, Phlox—Festuca), had only a small percentage of exposed bedrock at the surface (<5%); the other four were usually located on steeper slopes with 16-31% exposed bedrock. Phlox diffusa was dominant in five communities and codominated with Juniperus communis in the sixth.
The distribution and aerial extent of the Phlox communities reflected the influence of climate, substrate characteristics, and glacial history. The Phlox—Festuca community, for example, was limited to a narrow band from Hurricane Ridge southeast to Constance Pass. Sites supporting this grassy meadow type were dry but relatively stable with an average slope of 26° (Table A2). Exposed bedrock was low (2%) and plant cover was high (65%). The Phlox—Potentilla community occurred at higher elevations than the Phlox—Festuca type (1,864 m) often just below ridgecrests; these sites, characteristically blown free of snow in winter, were alpine rather than subalpine.
Four of the Phlox communities occurred mostly on exposed bedrock. Plants typically occurred within cracks and in depressions where soil had accumulated. Slope stability was higher than that of scree communities because exposed bedrock seemed to inhibit soil and rock movement—slope varied from 30 to 41°. Community types included Phlox—Carex phaeocephala, Juniperus—Phlox, and two phases of a Phlox—Pachistima type (one with Phacelia hastata). The Phlox—Carex phaeocephala type occurred only on Mount Dana and could have represented an early successional community or a steeper, wet-site version of the Phlox—Potentilla community. This community was unusual because wetter-site species, Carex spectabilis and Lupinus latifolius, codominated with the dry-site species, Phlox diffusa and Carex phaeocephala.
Carex spectabilis Communities
Environmental differences were not distinct among some of the plant communities at the wetter end of the classification scheme. Carex spectabilis dominated or was an important constituent of all communities in this group. The Carex—Valeriana and Vaccinium deliciosuin types were found on steep slopes (35-37°) from 1,615 to 1,675 m elevation with little bedrock at the surface (Table A2). The Carex—Lupinus type was widespread and occurred primarily on east to southeast-facing exposures averaging 1,800 m in elevation. The heath-dominated communities were dominated by Phyllodoce empetriformis on southerly exposures at elevations averaging 1,730 m and by Cassiope mertensiana on higher (1,830 m), northerly exposures.
There were differences among substrate characteristics of the five Carex spectabilis communities. Exposed bedrock averaged 17% in Cassiope communities compared to about 8.5% in Phyllodoce and Carex—Lupinus communities, and 2% or less in the Vaccinium and Carex—Valeriana types. Total rock cover was lowest in the Vaccinium and Phyllodoce communities.
Most plots classified in the Carex spectabilis or Phyllodoce community types were located in wetter areas of the park, west of the Elwha River and south of the Dosewallips River. One community, Vaccinium, was not observed or sampled in the dry Mount Angeles—Deer Park area. The Carex—Lupinus community was sampled in all areas of the park—it was more common on north-facing exposures in the Mount Angeles area as opposed to south-facing exposures in regions receiving greater precipitation. Some communities, such as the Phyllodoce empetriformis type, were more common in moderately sloped cirques and basins not sampled in this study (e.g., Seven Lakes Basin). These areas were not typical of summer range for goats (Stevens 1979).
Luetkea—Saxifraga Community
The Luetkea—Saxifraga type was sampled infrequently. It occurred on west to northwest-facing exposures—a fairly high proportion of the substrate was composed of larger rocks (11-70 cm). Plant cover was low (26%). This community occurred on sites that seemed to have been recently colonized by plants.
Interpretation
A comparison of communities described in this study with those delineated by Kuramoto and Bliss (1970) and Pfitsch and Bliss (1985) showed concurrence, but we identified additional community types. Differences among the studies arose because Kuramoto and Bliss (1970) concentrated on subalpine meadows; Pfitsch and Bliss (1985) examined communities in a comparatively small area; and the number of plots sampled in our study was greater (700+) than the ~40 sampled by Kuramoto and Bliss (1970) and the ~30 sampled by Pfitsch and Bliss (1985). Additionally, plant community patterns in goat summer range were often fine grained (i.e., community types often changed over distances of ≤5 m) because of interactions among substrate, time of snowmelt, slope stability, and precipitation.
Availability of soil moisture seemed to be the primary environmental factor controlling plant community composition and distribution. Average annual precipitation, date of snowmelt, aspect, and elevation combined to produce local moisture regimes. Previous investigators of Olympic Mountain subalpine and alpine communities reached similar conclusions (i.e., Fonda and Bliss 1969; Kuramoto and Bliss 1970; Canaday and Fonda 1974; Belsky and del Moral 1982; del Moral 1983b; Pfitsch and Bliss 1985; Smith and Henderson 1986).
Slope and substrate characteristics (i.e., particle size and the quantity of bedrock near the surface) played secondary but crucial roles in controlling plant community composition. The exact nature of interactions between substrate and available moisture remains unclear, but it seemed that sites with later snowmelt and bedrock near the surface developed heath communities while drier sites favored the Phlox—Pachistima community. Field observations suggested that parent material (i.e., sedimentary, slate, or basalt) also influenced plant community composition, although Kuramoto and Bliss (1970) reported similar chemistry among soils developed on different parent materials. Soil stability also affects subalpine and alpine vegetation patterns (Bliss 1969; Bell and Bliss 1973; Hansen 1976; Belsky and del Moral 1982; del Moral 1984; Pfitsch and Bliss 1985)—stability may be influenced by parent material and substrate size.
Plant community composition and distribution in the Olympic Mountains may reflect past glacial events. Kuramoto and Bliss (1970), for example, suggested that the Phlox—Festuca community could be a remnant of communities that were more widespread during maximum Holocene warmth (10,000-4,000 B.P.). The distributions of several plant species and communities suggest the existence of glacial refugia in the north and northeastern Olympic Mountains (see Chapter 3).
Subalpine and alpine plant communities are exposed to harsh environmental conditions. High winds, cold temperatures, snow, short growing seasons, freeze-thaw cycles, nutrient-poor soils, and soil moisture conditions play a role in community and species development (Bliss 1971). Microenvironmental factors such as soil temperature, wind, and soil moisture strongly influence plant growth and reproduction in these zones (e.g., Bliss 1956, 1971; Johnson and Billings 1962; Holway and Ward 1965; Billings 1974). These environmental constraints also influence subalpine and alpine meadow vegetation of the Olympic Mountains (Bliss 1969; Kuramoto and Bliss 1970; Canaday and Fonda 1974; Loneragan and del Moral 1984).
| <<< Previous | <<< Contents >>> | Next >>> |
appa2.htm
Last Updated: 12-Dec-2007