|
NATIONAL PARK SERVICE
Mountain Goats in Olympic National Park: Biology and Management of an Introduced Species |

|
| Mountain Goat Herbivory |
CHAPTER 10:
Responses of Vegetation to Reductions in Goat Density
E. G. Schreiner and A. Woodward
We established permanent plots in three areas of mountain goat summer range to evaluate plant responses to different levels of goat herbivory14 (Table 29). Each study area had relatively high initial goat density but differed in annual precipitation and the composition of plant communities. The three areas also differed physiographically: Klahhane Ridge extends east-west with mainly north and south exposures, Mount Dana is an isolated peak with all exposures, and Avalanche Canyon is a north-south valley with east- and west-facing exposures. We reduced each goat subpopulation and simultaneously measured vegetation to test the null hypothesis that reducing mountain goat density would not affect the relative abundance of plant species.
14Exclosures were not used because microenvironmental changes caused by fences may confound the effects of herbivore exclusion (Daubenmire 1940; Heady 1968). Additionally, fences in subalpine and alpine areas often trap snow (Schreiner 19987), which may profoundly affect plant growth.
Table 29. Location, sample size, and general information of areas
containing permanent plots.

(click on image for a PDF version)
Plots were placed in homogeneous vegetation where goat herbivory was initially high, and control plots were placed where herbivory was initially low. Plant cover and grazing levels were estimated on randomly located 1- x 1-m quadrats (n = 6-12) in each plot (Appendix A1). We sampled three times per year at Klahhane Ridge, two per year at Avalanche Canyon, and once per year at Mount Dana. More than one measurement per year was taken at Klahhane Ridge and Avalanche Canyon to ensure that we estimated plant cover at the seasonal maximum. Access to Mount Dana was too time-consuming to allow more than one sample per year.
We tested the null hypothesis by comparing total plant cover and cover of selected and nonselected plant species over time in plots with high and low initial levels of goat herbivory (declining herbivory versus stable or increasing herbivory at Avalanche Canyon; see below). We used regressions of plant cover on time and correlation coefficients (r) of cover with weather variables and goat density as descriptive statistics summarizing the association among the variables for each plot. Significant differences in these statistics among treatments were determined using a nonparainetric permutation test (Mehta et al. 1988).
Despite the large number of quadrats sampled (n = 172), the data set is still composed of small numbers of plots per treatment at any one site because we used mean cover to characterize vegetation. The minimum P-value attainable for a comparison of treatments with two replications (i.e., high initial level of goat use at Klahhane Ridge) and four replications (i.e., low initial level of goat use at Klahhane Ridge) was 0.0667, so we used 0.10 as the critical P-value. In the special instance where comparisons were between two treatments with two replications each, the minimum P-value was 0.1667, and we used 0.1667 as a significant difference, and we marked it with an asterisk (*).
Selected and nonselected forage species (Appendix A3) were determined separately for each area because forage chosen by mountain goats was known to be area specific (Chadwick 1983; Johnson 1983; see Chapter 9). There were 14 selected forage species and 16 nonselected forage species at Klahhane Ridge, 9 selected and 15 nonselected at Mount Dana, and no selected forage species at Avalanche Canyon. Goats foraged unselectively in Avalanche Canyon because most of the landscape was rock—cliffs, talus, or scree—hence there were relatively few plants to eat. We classified Avalanche Canyon plants as species grazed only for the first 2 years (n = 21) and species grazed throughout the study period or only during the last 3 years (n = 21). This classification provided groups whose response to grazing was expected to be analogous to the selected and nonselected categories.
Summer precipitation just before sampling was used as a measure of growing-season quality. July and August records from Port Angeles were used for Klahhane Ridge and Quilcene for Avalanche Canyon. June and July records from the Hoh Ranger Station were used for Mount Dana. We estimated Quilcene precipitation in 1992 with a regression based on Hoh Ranger Station data (r2 = 0.96) because Quilcene records were unavailable.
Trends in Plant Cover
Approximately one-third of total plant cover
( = 34.4%) was categorized as selected or nonselected by goats
at Klahhane Ridge. Mean cover of selected plant species was 9.0% and
29.7% for plots receiving (respectively) high and low initial levels of
goat use. In contrast, nonselected plant species cover was 11.3% in
plots with high levels of goat use and 11.0% in plots with low goat use.
As goat density was reduced, cover of selected plant species in areas
originally having high levels of goat use increased at a significantly
faster rate (i.e., greater slope of regression) than cover of selected
plant species in areas originally having low goat use (P =
0.0667; Fig. 36A; Table 30). Selected plant species cover also increased
faster than that of nonselected plant species in plots with high initial
levels of goat use (P = 0.1667* [special instance described in
third paragraph above]) but not in plots receiving low initial levels of
goat use (P = 0.3143). Total plant cover increased in plots with
high initial levels of goat use and decreased in plots with low initial
levels of use (P = 0.0667).
= 34.4%) was categorized as selected or nonselected by goats
at Klahhane Ridge. Mean cover of selected plant species was 9.0% and
29.7% for plots receiving (respectively) high and low initial levels of
goat use. In contrast, nonselected plant species cover was 11.3% in
plots with high levels of goat use and 11.0% in plots with low goat use.
As goat density was reduced, cover of selected plant species in areas
originally having high levels of goat use increased at a significantly
faster rate (i.e., greater slope of regression) than cover of selected
plant species in areas originally having low goat use (P =
0.0667; Fig. 36A; Table 30). Selected plant species cover also increased
faster than that of nonselected plant species in plots with high initial
levels of goat use (P = 0.1667* [special instance described in
third paragraph above]) but not in plots receiving low initial levels of
goat use (P = 0.3143). Total plant cover increased in plots with
high initial levels of goat use and decreased in plots with low initial
levels of use (P = 0.0667).
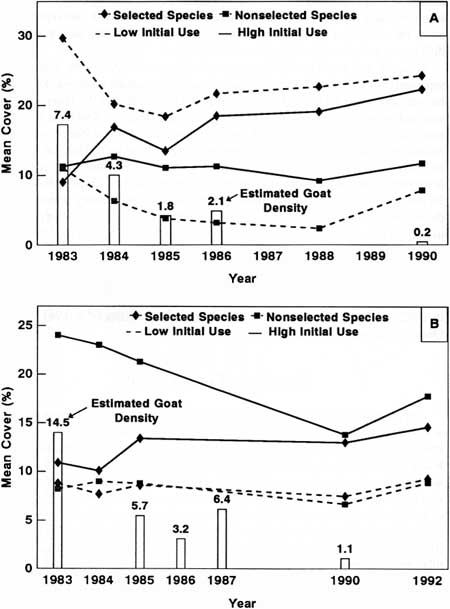
|
| Fig. 36. Estimated mountain goat density and mean cover of selected and nonselected goat forage species in plots with high versus those with low initial levels of goat use at Klahhane Ridge (A) and Mount Dana (B). |
Table 30. Mean slope of regression (b) of mean plant cover over time and mean correlation coefficients (r) of mean cover with estimated mountain goat (Oreamnos americanus) density. Goat density estimates in 1983 and 1990 did not correspond with 1988-1992 vegetation sampling in Avalanche Canyon, so the correlations between plant cover and goat density could not be determined.
| Klahhane Ridge | Mount Dana | |||
| Treatment Species group |
Slope (b) |
Correlation with goat density (r) |
Slope (b) |
Correlation with goat density (r) |
| Low initial goat use | ||||
| Selected speciesa | -0.25b | 0.50c | 0.04 | 0.17 |
| Nonselected species | -0.31 | 0.23 | -0.07 | -0.20 |
| Total cover | -0.42d | 0.53e | -0.87 | 0.39 |
| High initial goat use | ||||
| Selected species | 1.56b,j | -0.84c | 0.45f,g | -0.69h,i |
| Nonselected species | -0.15j | -0.17 | -1.02f | 0.53h |
| Total cover | 2.80d | -0.87e | -0.61g | 0.50i |
| Avalanche Canyon | ||||
| Treatment Species group |
Slope (b) | |||
| Declining use | ||||
| Grazed first 2 years | 1.82k | |||
| Grazed throughout | 0.24k,l | |||
| Total cover | 4.35 | |||
| Stable or increasing use | ||||
| Grazed first 2 years | 1.02 | |||
| Grazed throughout | 2.00 | |||
| Total cover | 4.04 | |||
aNo statistical comparisons were made with selected species in low intitial goat use plots at Mount Dana because sample size was too small. b,c,iSignificantly different at P ≤ 0.10 using a nonparametric permutation test (Mehta et al. 1988). j,k,lSignificantly different at P ≤ 0.17 (permutation test)—this was the minimum possible P-value given the sample size. | ||||
Mean cover of selected plant species increased (10.9-14.6%) as goat density was reduced in plots receiving high initial levels of goat use at Mount Dana (Fig. 36B). This contrasted significantly with a decrease in cover of nonselected plant species (24.0-17.8%; P = 0.0970) and total plant cover (42.1-40.1%; P = 0.0482) in these plots (Table 30). We did not compare cover of selected plant species in plots with high initial use versus low initial use because sample size was too small for statistical tests (i.e., selected plant species occurred in only one of two plots with low initial levels of goat use). Total plant cover and cover of nonselected plant species were not significantly different between those plots with high and those with low initial goat use.
Avalanche Canyon data were compared using plots with decreasing goat use versus plots with stable or increasing goat use (Fig. 37) because all plots had high initial levels of goat use (i.e., there were no areas with low goat use). We compared species grazed only in the first 2 years of the study with those that were grazed throughout or only during the last 3 years of the study. Species grazed initially in the study were expected to increase, and those grazed later were expected to decline. Plant cover of species grazed only the first 2 years increased faster than species grazed throughout or late in the study (P = 0.1667*) in plots experiencing declining goat use. These two groups were not significantly different from each other in plots with stable or increasing use.
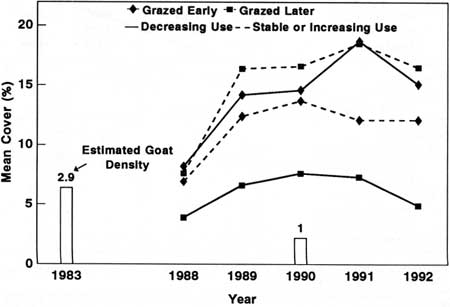
|
| Fig. 37. Estimated mountain goat density and mean cover of plants grazed early (1988-89) and later (1988-92 or 1990-92) in plots with decreasing versus those with stable or increasing levels of goat use at Avalanche Canyon. |
Goat Density, Precipitation, and Plant Cover
Cover of selected plant species was negatively correlated with goat density in plots receiving high initial levels of goat use at Klahhane Ridge and Mount Dana (Table 30). The correlation coefficients between selected plant species cover and goat density in plots with high versus low initial goat use were significantly different at Klahhane Ridge (P = 0.0667)—sample size was too small to perform the test on data from Mount Dana. The correlations of nonselected plant species cover and selected plant species cover with goat density were significantly different at Mount Dana (P = 0.0188) but not on Klahhane Ridge. There were no differences between the correlations of nonselected plant species cover and goat density in high versus low initial use plots at Klahhane or Dana. Goat density estimates in the Avalanche Canyon area (1983, 1990) did not correspond with years of vegetation sampling (1988-92) so the association between goat density and plant cover could not be determined.
Total plant cover in plots receiving high initial levels of goat use was negatively correlated with goat density and positively correlated with density in plots with low initial levels of goat use on Klahhane Ridge (P = 0.0667). The correlation between goat density and total cover at Mount Dana was positive in high and low initial-use plots. Correlations using Avalanche Canyon plots were not possible (see above).
Correlation coefficients between summer precipitation and plant cover were not significantly different among treatments within study areas (Table 31). Correlations between precipitation and cover at Mount Dana were not consistent among treatments—two were positive and four were negative. Summer precipitation at the Hoh River (34 km distant at low elevation) may not have reflected growing season quality for Mount Dana because mean cover declined substantially in 1990 even though summer rainfall that year was the highest during the study.
Table 31. Correlation coefficients (r) between mean plant cover and summer precipitation just before the sample period.
| Treatment Species group |
Klahhane Ridge (1983-90) | Mount Dana (1984-92) |
| Low initial goata use | ||
| Selected species | 0.32 | -0.37 |
| Nonselected species | 0.29 | 0.02 |
| Total cover | 0.61 | -0.02 |
| High initial goat use | ||
| Selected species | 0.14 | 0.13 |
| Nonselected species | 0.39 | -0.61 |
| Total cover | 0.25 | -0.31 |
| Avalanche Canyon (1988-92) | ||
| Declining goata use | ||
| Grazed first 2 years | 0.82 | |
| Grazed throughout | 0.46 | |
| Total cover | 0.48 | |
| Stable or increasing goat use | ||
| Grazed first 2 years | 0.44 | |
| Grazed throughout | 0.54 | |
| Total cover | 0.58 | |
aMountain goat (Oreamnos americanus). | ||
Plant Community Responses
The plots used to study vegetation response to lowered mountain goat density represented seven plant communities. Klahhane Ridge plots (6) were all classified as Phlox—Festuca (one also fit the unstable herb-meadow community of Pfitsch and Bliss 1985). Each plot at Mount Dana (5) represented a different community (Phlox—Carex phaeocephala, Phlox—Festuca, Phlox—Pachistima, Carex spectabilis—Lupinus latifolius, and Phyllodoce empetriformis) and four communities were represented by the six plots in Avalanche Canyon (Phlox—Pachistima, one; Phlox—Festuca, one; Phacelia—scree, three; and Delphinium—scree, one). Community types were replicated within each treatment at Klahhane Ridge but not at Mount Dana or Avalanche Canyon. Consequently statistical analyses were possible only with Klahhane Ridge data.
Festuca idahoensis and Achillea millefolium were major constituents of the plant communities at Klahhane Ridge. They have a competitive relation mediated by herbivory—that is, A. millefolium dominated when F. idahoensis was grazed, and Festuca dominated when neither was grazed (del Moral 1985). Festuca idahoensis increased in plots where goat use was originally high and decreased in plots where use was originally low (significantly different slopes at P = 0.0667; Fig. 38). Achillea millefolium decreased in plots with high initial goat use but not in low initial use plots (P = 0.0667). Also, percent cover of A. millefolium was positively correlated with goat density in high use plots, which differed from correlations in low use plots (P = 0.1667*). This contrasted with the negative relation between F. idahoensis cover and goat density in plots receiving high initial goat use. Correlation coefficients between F. idahoensis cover and goat density in high versus low use plots also were significantly different (P = 0.0667).
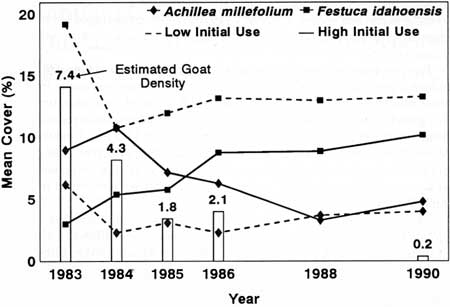
|
| Fig. 38. Estimated mountain goat density and mean cover of Achillea millefolium and Festuca idahoensis in plots with high versus those with low initial levels of goat use at Klahhane Ridge. |
The relative abundance of plant species at Klahhane Ridge changed over time in plots receiving high initial goat use but not in those receiving low initial use. For example, the disturbance-oriented A. millefolium dominated a high initial use plot (~10% cover) in 1983 and Cirsium edule and Phlox diffusa codominated with about 6% cover each. By 1990, Cirsium and Phlox shared dominance (~9% cover) and F. idahoensis had ~7% cover; Achillea had decreased to about 6%. A similar pattern of change occurred in the other plot with high initial use, but different species were involved. Relative cover among species in the four plots with low initial goat use remained essentially constant (Schreiner et al. 1993). Plant relative abundance in the high use plots seemed to be approaching that of the low use plots toward the end of the study.
We noted similar trends in behavior among several plant species at Mount Dana across community types (Schreiner et al. 1993). For example, three selected forage species (Carex spectabilis, Juncus parryi, and Aster paucicapitatus) occurred in three different communities and each increased in mean cover. By contrast, the disturbance-oriented Lupinus latifolius decreased by half or more in two high initial-use communities where it occurred. These changes modified relative abundance among codominant species in the three communities that received high initial levels of goat use. Similar changes did not occur in the two communities receiving low initial levels of goat use. Relative abundance of species in Avalanche Canyon did not change consistently within treatments or species groups.
Interpretation
Our results demonstrated that subalpine and alpine vegetation responded to lowered goat density in both the climatically wet and dry regions of Olympic National Park. As groups, selected goat forage species increased and nonselected forage species decreased as goat density was reduced. The selected and nonselected goat forage species groups (46 species total) each included bunchgrasses, shrubs, and taprooted forbs as well as rhizomatous forbs, grasses, and sedges. Caughley (1970a) reported analogous changes in plant cover following introduction of a mountain ungulate but for only eight forage species.
The reduction in goat density at Avalanche Canyon was insufficient to induce a pronounced response in vegetation, in contrast to results from the other two study areas. Goat density was only reduced from an estimated 3 to 1 goat/km2 in the Avalanche Canyon area. This compares to reductions of from 7 to 0.2 goat/km2 at Klahhane Ridge and from 14.5 to 1.1 goats/km2 at Mount Dana. Additionally, there was evidence that effective goat density did not change in Avalanche Canyon. Five goats grazed in permanent plots as we sampled them in 1992, even though 12 goats were removed from the general area during 1988-89.
Subalpine and alpine vegetation of the Olympic Mountains is characterized by plant communities shaped by soil moisture availability and level of disturbance (Belsky and del Moral 1982; del Moral 1983b; Pfitsch and Bliss 1985; Appendix A2). Our results confirm the importance of disturbance (i.e., herbivory) in controlling the relative abundance of plant species at high elevations (>1,520 m). Individual disturbance-oriented plant species decreased and selected plant species increased as levels of disturbance (goat density) were reduced. These changes took place in sparsely vegetated plant communities on unstable substrates as well as those dominated by shrubs, forbs, and sedges on comparatively stable substrates. We interpret this to mean that goats caused increased disturbance in a wide variety of plant communities, irrespective of species composition. The null hypothesis that plant relative abundance would not respond to changes in mountain goat density was rejected.
The effects demonstrated here of goats on vegetation occurred mainly on summer ranges where resources (i.e., plants) were unlikely to limit the population (Pfitsch and Bliss 1985; Houston and Stevens 1988). Goat effects on winter range, especially on cliffs and rock outcrops where plant cover is extremely low, could have been even more dramatic.
| <<< Previous | <<< Contents >>> | Next >>> |
chap10.htm
Last Updated: 12-Dec-2007