
|
Texas Bureau of Economic Geology
Guide to the Permian Reef Geology Trail, McKittrick Canyon, Guadalupe Mountains National Park, Texas |
REEF
Brenda L. Kirkland, Susan A. Longacre, and Emily L. Stoudt
The next portion of the Permian Reef Geology Trail cuts through ~152 m (~500 ft) of reef facies—the massive Capitan Formation (Richardson, 1904; King, 1942, 1948). At an elevation of ~1,905 m (~6,250 ft) (figs. 2, 7, and 20), the in-place reef facies interfingers with forereef debris. The reef facies extends up to ~2,060 m (~6,750 ft) where reef boundstones are interbedded with outer-shelf grainstones. The presence of the fusulinid Polydiexodina along part of the trail and stratigraphic correlations up-canyon indicate that the Capitan Formation along the trail is equivalent to the middle and upper Yates Formation (Yates B and C of Newell and others, 1953) (fig. 4). This part of the reef is older and stratigraphically below the Tansill-equivalent reef facies frequently visited in Walnut and Dark Canyons located 43 km (27 mi) and 55 km (34 mi), respectively, to the northeast.
Before considering features present in the reef facies of the Capitan Formation, it is helpful to review some fundamental aspects of reefs. A modern reef community consists of a diverse array of organisms, many with calcareous skeletons, living from sea level down to a depth of ~91 m (~300 ft) (James, 1983). Essential components of a modern reef can be categorized into four groups. First is the community of frame-building and binding organisms that form the structure of the reef. Second is marine cement, which, along with binding organisms, lithifies the reef. Third is the group of bio-eroders that, by their activities, break down the reef and create the fourth component, internal sediment. Along with marine cement, internal sediment can fill any available space within the reef structure (Ginsburg and Lowenstam, 1958; Moore, 1989; Tucker and Wright, 1990). Evidence of most of these components is displayed in the stops along this part of the trail.
In contrast with the transported blocks of Tansill-equivalent reef that occur on the slope, reefal boundstones along this part of the trail are preserved in place. Compared with the Tansill-equivalent reef exposed at the mouth of Walnut Canyon and in the slope blocks at McKittrick Canyon, the Yates-equivalent reef along the Permian Reef Geology Trail exhibits a greater diversity of sponges, less Archaeolithoporella, and fewer and smaller fans of botryoidal cement. Also in contrast to the slope section is the apparent lack of both disseminated quartz silt and interbedded siltstones through the segment of the reef exposed by the trail.
Debate about depth of water above the living Permian reef continues. As noted in the introduction, the depositional profile for Capitan sediments varied vertically and laterally through time. Yurewicz (1977), Babcock (1977), and Toomey and Babcock (1983) suggested that the Capitan reef accumulated in progressively shallower water from the time of deposition of the Seven Rivers Formation to that of the Tansill Formation. Babcock (1977) interpreted the presence of Collenella, abundance of algal and other shallow-water elements, and biotic diversification in the youngest, shelfward facies of the reef in Walnut and Dark Canyons as indicative of shallower water depths. Collenella is present in the uppermost outcrops of the Yates-equivalent Capitan in McKittrick Canyon (Stop 22), suggesting a similar shallowing of the Yates-equivalent reef. The abundance of framework and binding components suggests that the reef could resist waves, but water depth above the reef cannot be estimated from the reef unit alone.
As discussed in the next section of the trail guide, the geometry of the outer-shelf beds suggests "fall-in" toward the equivalent reef (figs. 5a, c and 43). The fall-in bed geometry suggests that the reef was deposited downdip from the highest part of the shelf profile, in water depths from 30 to 43 m (100 to 140 ft) during Seven Rivers deposition (Yurewicz, 1977; Hurley, 1978) and in depths as shallow as 9 to 15 m (30 to 50 ft) during deposition of the Yates-B-equivalent reef (Kerans and Harris, this volume, p. 32). Different conclusions were reached in a sedimentologic study of the upper Capitan and Tansill Formations by Kirkland-George (1992), who proposed that the Tansill-equivalent reef grew to sea level and protected a shallow-water lagoon on the outer shelf (fig. 5d).
Key features within the reef portion of the trail include organisms, cements, and internal sediments. The Guadalupian-aged fauna of the massive Capitan is diverse, but it is dominated by a variety of calcareous sponges and bryozoans, rather than corals as in modern reefs. Volumetrically the most important binding element of the reef is the rhodophyte Archaeolithoporella, which occurs along with prodigious amounts of marine botryoidal-fan cement (fig. 27a). Other marine cements include isopachous rims of calcite that are visible without a hand lens (Stop 21). Macroscopic examples of layered internal sediment filling both framework voids and possible neptunian dikes are abundant along the trail (Stop 19).
Diagenetic events recorded in the Capitan reef will be discussed in greater detail at Stop 21, but note that typical cements that are present along the trail are dominantly (1) marine botryoidal fans, (2) marine isopachous fibrous crusts, and (3) several generations of calcspar.
STOP 15. Interbedded Basal Reef and Slope
Deposits
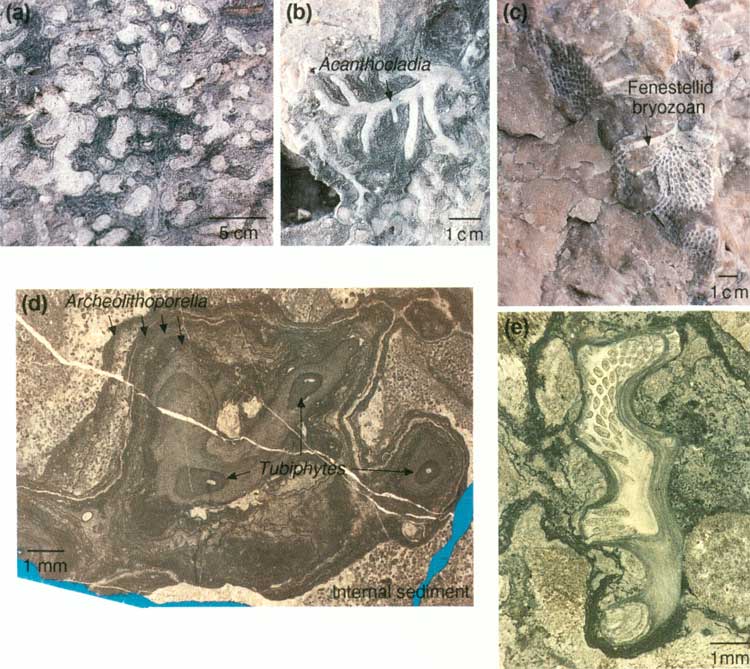
Stop 15 includes a series of locations beginning a ~1,905 m (~6,250 ft), at the first rock exposure beyond the grassy, scree-covered slope below the steep walls of the massive reef. This stop extends for 142 m (465 ft) along the trail to just around switchback D (figs. 2 and 20) in the saddle east of the massive reef outcrop. A large cephalopod and a mass of Tubiphytes (a tiny, porcellaneous, problematic organism that weathers whitish on outcrop) are displayed in the first major exposure along this part of the trail. Twenty-seven meters (90 ft) farther up the trail is a reddish-weathering recess (fig. 27a) that shows the typical reef boundstone of sponges and bryozoans encrusted with Archaeolithoporella, botryoidal-fan cements, and Tubiphytes. Laminated internal sediment also occurs in reef cavities. A variety of ramose and fenestellid bryozoans, including Acanthocladia, are exposed in the floor and north side of the trail just below switchback D (figs. 20 and 27b). Several cephalopods (fig. 27c) are present in the trail ~3 m (~9 ft) beyond the 6,300-ft marker just above switchback D.

|
| FIGURE 27. Stop 15, basal reef: (a) typical reef boundstone with interlaminated Archaeolithoporella (dark-gray thin lines) and rust-colored botryoidal-fan cement encrusting sponges; some voids filled with internal sediment (1/4 actual size), (b) bryozoans in floor of trail (1/2 actual size), (c) cephalopod in wall of trail (1/4 actual size), (d) view looking south of reef trail showing locations of switchbacks D through F, Stops 16-19, and an approximate time line through the reef derived from plotting the youngest occurrence of the fusulinid Polydiexodina. In the reef exposures along the trail, this fusulinid has been found only west (right) of this red line. (click on image for an enlargement in a new window) |
Interbedded with the reefal boundstone immediately below switchback D are steeply dipping, Yates-C-equivalent skeletal grainstones and packstones deposited in the transitional reef/slope environment. The bedded packstones and grainstones exposed at switchback D on the east side of the saddle are Tansill-equivalent slope deposits.
STOP 16. Well-Exposed
Sponge-Archaeolithoporella Boundstone
Stop 16, which straddles the trail ~64 m (~210 ft) beyond switchback D (fig. 27d), is another excellent exposure of typical Yates-equivalent Capitan reef framework. The three common elements of the Capitan reef—organisms, marine cement, and laminated internal sediment—are all well displayed in this exposure. The more common sponges occurring throughout the reef are illustrated in figure 28. Based on a census of the sponges along the trail, the most common sponge is an elongate form with geometrically regular pores that represents Lemonia cylindrica (formerly Guadalupia cylindrica); both small and large forms of this sponge are found in almost all localities. Other sponges mixed with this form are Guadalupia, Amblysiphonella, Cystauletes, Girtycoelia, and Cystothalamia. Almost all reef outcrops also contain several growth forms of delicate to robust ramose and fenestellid bryozoans. With the exception of the transition into the shelf sediments (Stop 22), neither spatial nor temporal zonation was detected in the fauna. The lack of zonation can be accounted for by the filter feeding habits of most of the organisms that compose this Permian reef; in contrast to modern corals, they were not constrained by the occurrence of light-sensitive zooxanthellae.

|
| FIGURE 28. Stop 16, sponges of the Capitan reef. Photographs are almost 1/2 actual size. (click on image for an enlargement in a new window) |
STOP 17. Tubiphytes/Acanthocladia
Boundstone
Stop 17, ~46 m (~150 ft) beyond Stop 16 (figs. 27d and 34), contains a large mass of Tubiphytes/Acanthocladia boundstone. The 6,400-ft marker is ~100 m (~300 ft) beyond this exposure; that marker is 46 m (150 ft) from switchback E. At Stop 17, typical Acanthocladia fronds are well displayed in a whitish specimen located 0.6 m (2 ft) above and to the left of the trail (fig. 29b and 29e). Although encrusting Tubiphytes is common throughout the reef, it is particularly well-developed here (fig. 29a) along with fenestellid bryozoans (fig. 29c). Some organisms appear to be preserved in growth position, although others are debris of probably broken specimens. Voids between the branches of the organisms are filled with isopachous, fibrous, radiaxial cement and laminated internal sediment (fig. 29d). Although the best exposure along the trail measures only 2 by 3 m (6.6 by 10 ft), the mass extends for at least 25 m (75 ft) along and above the trail.
|
| FIGURE 29. Stop 17, Tubiphytes and Acanthocladia in reef: (a) Tubiphytes colony (1/6 actual size), (b) branching Acanthocladia (1/2 actual size), (c) fenestellid bryozoan exposed in trail (1/4 actual size), (d) photomicrograph of Tubiphytes colony encrusted by Archaeolithoporella, (e) photomicrograph of cross section of Acanthocladia. (click on image for an enlargement in a new window) |
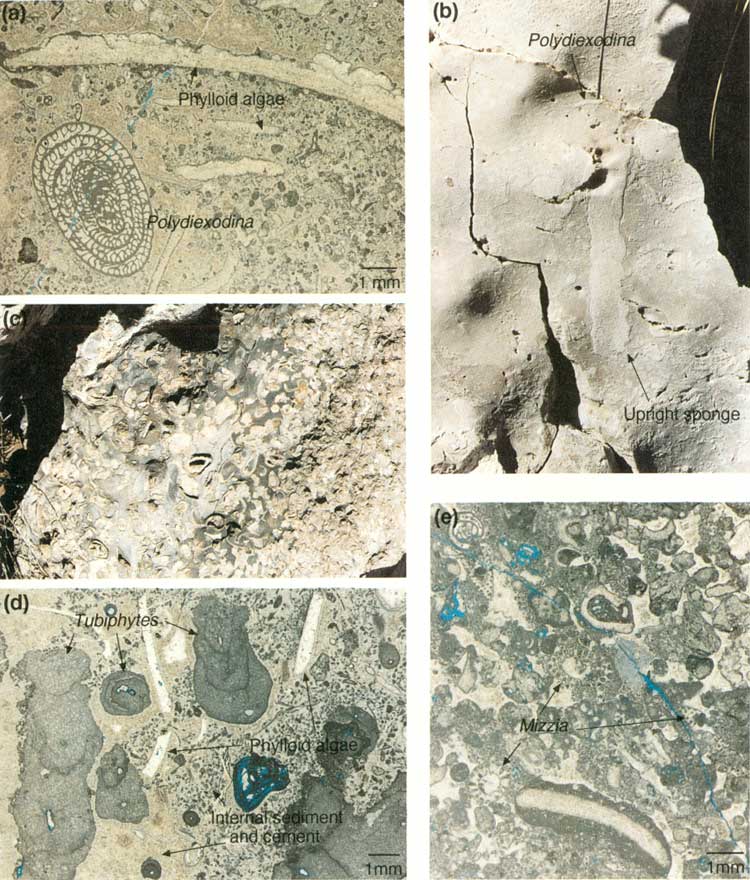
STOP 18. Fusulinids within the Reef
Stop 18 is located at switchback F (figs. 27d and 34) where the large, age-diagnostic fusulinid, Polydiexodina (fig. 30a), occurs among upright sponges (fig. 30b). Polydiexodina is 25 to 38 mm (1 to 1.5 inches) long, easily seen, and characteristic of the Seven Rivers to Yates B (middle Yates) interval in the shelf facies (Newell and others, 1953). Observed occurrences of Polydiexodina are on the north side of the line drawn on figure 27d, making this portion of the reef equivalent to the Yates B. Its absence between the line and the base of the Tansill-equivalent slope deposits (switchback D, fig. 20) suggests that the reef interval along this part of the trail is equivalent to the younger Yates C. Note that the slope of the line, which may approximate a time line, is near vertical, suggesting a rather steep-fronted reef profile.
|
| FIGURE 30. Stop 18, fusulinids in reef, and Stop 19, pockets or neptunian dikes: (a) photomicrograph of fusulinid Polydiexodina and phylloid algal fronds, (b) outcrop photo of upright sponge and Polydiexodina (1/4 actual size), (c) outcrop expression of a neptunian dike containing large brachiopod shells (1/5 actual size), (d) photomicrograph of Tubiphytes and phylloid algal fragments encased in internal sediment and marine cement from dike site 2, (e) photomicrograph of Mizzia and other backreef skeletal grains observed at third dike or pocket in reef. (click on image for an enlargement in a new window) |
The trail crosses the Yates B/C boundary twice more before reaching Stop 19, ~126 m (~415 ft) up the trail beyond switchback F (fig. 27d). Along the trail are several pockets filled with yellowish-orange-stained, silt-sized dolomite. As discussed in the slope section of the trail guide, similarly colored layers in the slope are composed of shelf-derived quartz siltstone and very fine grained sandstone. Although similar sandstone layers may occur within the Yates-equivalent reef, they are not obvious along the trail.
STOP 19. Pockets or Neptunian Dikes within the
Reef
Stop 19 includes a series of locations along 78 m (255 ft) of trail (figs. 27d), beginning ~21 m (~69 ft) beyond the 6,500-ft marker. At least three sites along this part of the trail contain organisms that either are markedly different from the typical reef organisms or are badly broken and clearly not in place. These assemblages may represent (1) fill within neptunian dikes that cracked open during reef growth and later were infilled by allochthonous grains, (2) pockets within the reef that were infilled with allochthonous debris, or (3) anomalous microcommunities within the reef.
The first and most striking of these sites predominantly contains large brachiopods (fig. 30c) but also contains gastropods and Polydiexodina. This location displays a distinct size zonation from coarser material in the center of the deposit to finer material along the margin. Two cephalopods occur along the trail between the first and second sites of Stop 19. The second site, ~23 m (~75 ft) farther up the trail, is dominated by fragmented Tubiphytes (fig. 30d), scattered plates of phylloid algae, sponges, and Polydiexodina. The grains are cemented with isopachous marine cement. Very small peloids (probably internal sediment) are surrounded by isopachous rims of marine cement. The third site, 53 m (174 ft) beyond site 2, contains a distinctive assemblage of Polydiexodina, other foraminifers, Mizzia (a green alga common in shelf deposits), phylloid algal plates, echinoderm and brachiopod fragments, and intraclasts/oncolites (fig. 30e). Most of these grains are cemented by thin rims of isopachous marine cement.
Overall, these pockets or dikes appear to coincide with the location of Tertiary fracture sets that are clearly visible when the trail is viewed from Stop 20. Note that the fractures virtually parallel the line marking the youngest occurrence of Polydiexodina in figure 27d. These Tertiary fractures may represent reactivation associated with Permian neptunian dikes or they may coincide with "growth surfaces" on the reef face.
STOP 20. Typical Reef Outcrops and Cave
Fill
Stop 20 incorporates ~46 m (~150 ft) of trail (figs. 27d and 34). The outcrops along this stop display particularly well-developed layers of internal sediment (fig. 31a) within reef boundstone. However, the most interesting feature of this portion of the trail is a small outcrop of cave fill (fig. 31b) preserved about 5 m (15 ft) down trail from the ridge point. This relict cave is filled by probable waterlaid floor sediments (fig. 31b) consisting of brownish-yellow carbonate containing peloids and ostracodes, interbedded with calcite rafts (fig. 31c) (Chafetz and Butler, 1980; Folk and others, 1985). Directly above these deposits is an exposure of banded travertine consisting of elongate columnar blades of light-tan calcite (fig. 31b). Petrographically similar travertine fills many Tertiary-age fractures throughout the Guadalupe Mountains.

|
| FIGURE 31. Stop 20, Permian internal sediment and cave fill in reef: (a) outcrop of layered internal sediment overlain by marine-cemented reef boundstone containing sponges and Archaeolithoporella (1/5 actual size), (b) outcrop exposure of small collapsed cave with layered, iron-stained sediment fill (f), calcite rafts (r), and honeycomb-weathered travertine (t), (c) photomicrograph of cave sediment fill containing peloids, ostracodes, and calcite rafts, plane light, (d) isotopic data from principal reef cements, dolomite, and speleothems (from Given and Lohmann, 1986; Hill, 1987; Mruk, 1989; Melim, 1991). Dolomite values plotted as average ± standard deviation; arrow indicates shelfward trend toward heavier oxygen isotopes, suggesting increasing salinity of dolomitizing fluids (Melim, 1991). (click on image for an enlargement in a new window) |
D. Mruk (personal communication, 1992) measured carbon and oxygen isotopes from the brownish-yellow sediments, calcite rafts, and travertine (fig. 31d). Her data fall in the same range as that of similar Quaternary calcite rafts from Carlsbad Caverns described by Hill (1987). When this small cave formed is uncertain; it may have developed concurrently with numerous other caves in the Guadalupe Mountains. However, the oxygen isotope values fall in the same range as spar I (see next stop), whose time of precipitation was most probably Permian.
STOP 21. Reef Diagenesis
Stop 21 begins 46 m (150 ft) beyond the 6,600-ft marker (fig. 34) in a distinctive, fresh cliff-face and extends for 91 m (300 ft) along the trail. The complex diagenetic sequence in the Capitan reef, as described in detail by Mruk (1985, 1989), Crysdale (1986), and Melim (1991), is well-developed. Many of the cements (fig. 32a) are visible on outcrop; others require examination of thin sections in plane light, or in some cases cathode luminescence, for recognition.

|
| FIGURE 32. Stop 21, reef cement: (a) schematic diagram of cement types in reef (modified from Mruk, 1989), (b) botryoidal-fan cement (note relict aragonite needle morphology) and encrusting layers of Archaeolithoporella (A). Aragonite alteration varies from almost unaltered (1) to replaced by sparry calcite (S) to dissolved and/or replaced by dolomite (4), (c) outcrop expression of major cement types (1/5 actual size), (d) photomicrograph of major cement types, (e) outcrop expression of calcspars and marine cements (1/5 actual size); coarse-crystalline milky spars are primarily spar II, (f) outcrop photo of light- and dark-colored botryoidal fans (1/5 actual size). (click on image for an enlargement in a new window) |
The earliest-formed diagenetic components are dark-gray and light-brown botryoidal fans (cement 1) filling framework voids within the reef (fig. 32a-32f); the botryoids are large 1.5 to 5 cm (0.6 to 2 inches) and interlaminated commonly with the alga Archaeolithoporella. Petrographic and isotopic evidence (figs. 31d and 32b) suggests an aragonitic precursor and a marine origin (Newell, 1955; Loucks and Folk, 1976; Mazzullo and Cys, 1978; Given and Lohmann, 1985, 1986; Mruk, 1985, 1989). A second type of marine cement, which is smaller but also visible on outcrop, is an isopachous cement that encrusts surfaces of botryoids and lines framework voids (cement 2; fig. 32a). These isopachous rims are thin (0.5-4 mm) with up to four layers visible on outcrop (fig. 32c and 32e). In thin section, isopachous rims were followed by inclusion-rich prismatic cement (Mruk, 1989) (cement 3; figs. 32d and 33a). In many cases, cements 2 and 3 have radiaxial extinction patterns. Morphology and isotopic signatures (fig. 31d) of these cements indicate formation from marine fluids.
Mruk (1989), Melim and Scholle (1989), and Melim (1991) recognized significant dolomitization, dissolution, fracturing, and brecciation events after marine cementation and before precipitation of the first calcspar cements. Scattered dolomite crystals also postdate calcspar 1 (fig. 33b and 33c). Although the Capitan reef is not as extensively dolomitized in the Guadalupe Mountains as it is in the subsurface (Garber and others, 1989), dolomite is present in varying amounts in the reef and slope deposits. In addition to pervasive dolomitization along fractured and brecciated zones described at Stop 22, petrographic examination reveals scattered small (0.2 mm) dolomite rhombs and leached rhombic pores (fig. 33c).

|
| FIGURE 33. Stop 21, reef cement, and Stop 22, Collenella zone; (a) photomicrograph of cathodoluminescence patterns in spar I (zoned dull to bright), spar II (zoned mostly bright), and spar III (unzoned dull), (b) dolomite cement 4 capping spar I and marine cements, (c) partial dissolution of dolimite 4, dissolved rhombs infilled with blue epoxy, (d) photomicrograph of luminescent spar II showing corroded crystal margins, (e) Collenella head, shown 2/3 actual size, (f) upper reef displaying dolomitization (light cream color) associated with fractures (lens cap for scale). (click on image for an enlargement in a new window) |
Three generations of spar are present in the reef; each is distinguished by cathodoluminescence and isotopic composition (Mruk, 1989) (figs. 31d and 33a). The first and third generations are distinguished only by luminescence in thin section. Spar I (using the terminology of Given and Lohmann, 1986, and Mruk, 1989) consists of small crystals (0.25 to 0.5 mm) formed in optical continuity with the inclusion-rich prismatic cement (fig. 32d). Spar II, particularly abundant along this portion of the trail (fig. 32c and 32e), formed as much larger crystals (0.5 to 10's of cm [0.6 to 4 inches]) and composes 10 percent of the cement in the massive facies (Mruk, 1989). Spar II probably formed during Mesozoic burial of the Capitan carbonates, as indicated by isotopic signatures and two-phase fluid-inclusion studies (Crysdale, 1986; Mruk, 1989). An episode of dissolution and corrosion (fig. 33d) preceded precipitation of spar III (Mruk, 1989) on and within corroded spar I crystals. Spar III is distinguished by dull luminescence (fig. 33a). Vugs filled with calcite spar are prominent along the trail; however, cores from the subsurface reef display similar vugs that are filled with anhydrite and/or gypsum (Garber and others, 1989). Melim and Scholle (1989) suggested that evaporites originally filled vugs in the Guadalupe Mountains but later were replaced by calcite spars II and III.
STOP 22. Upper Reef— Collenella, Fusulinids, and Dolomite
Stop 22 occurs ~18 m (~60 ft) below switchback G (fig. 34). The fusulinid Polydiexodina is present, indicating that this highest reef exposure along the trail is no younger than Yates B. The fauna is dominated by Collenella (a problematic alga that occurs in upper reef facies in other canyons) (fig. 33e), Acanthocladia, and partly articulated crinoids; the diverse fauna also includes corals, gastropods, bivalves, cephalopods, several types of sponges, and Archaeolithoporella. The presence of Collenella here but not elsewhere within the reef section suggests that water depths may have shallowed during deposition of the Yates B. Kerans and Harris (this report) describe evidence of exposure of the outer-shelf facies at the top of Yates B (Stop 24). Although water level probably did not drop below the top of the reef at this time, the presence of Collenella and the diverse fauna may reflect the shallowing or fall of relative sea level that resulted in exposure further up on the shelf. Shelf grainstones and reef boundstones are interbedded just below switchback G.

|
| FIGURE 34. Oblique aerial photograph of upper portion of Permian Reef Geology Trail showing reef Stops 17-22 and shelf Stops 23-28, elevation markers, major switchbacks, and key stratigraphic relations. Stop 24 details relations associated with the lower sequence boundary shown, Stop 26 covers key sandstone marker beds used to define the top of the Yates Formation, and Stop 28 illustrates facies changes reflecting maximum flooding within the uppermost (Tansill 1) sequence. (click on image for an enlargement in a new window) |
A brecciated zone occurs near the reef/outer-shelf transition on both sides of the nearby spur where the trail crosses it twice, just below and above switchback G. During shallow burial, the rigid, marine-cemented reef fractured, probably due to compaction of the underlying, unconsolidated slope facies. Dolomitization altered the rock along fracture walls, creating dolomite halos (fig. 33f). Dolomite is also present in permeable beds in the slope, with the amount of dolomitization decreasing downslope.
Average carbon and oxygen stable isotope values for fabric-preserving dolomite are increasingly heavy shelfward of the reef (fig. 31d), suggesting that the salinity of the dolomitizing fluid was increasing across the shelf (Melim, 1991). On the basis of (1) isotopic values, (2) distribution of dolomite (decreasing basinward), and (3) petrographic evidence for timing of dolomitization, Melim (1991) attributed dolomitization to refluxing of Permian saline fluids from the shelf down into the reef and slope along fractures and permeable zones, similar to the model of Adams and Rhodes (1960).
Summary
The massive Capitan reef along the trail exposes a diverse marine fauna dominated by a variety of calcareous sponges and several forms of bryozoans and algae. Little biological zonation is apparent within the reef along the trail. As indicated by the distribution of the fusulinid Polydiexodina, the trail crosses reef facies equivalent to both Yates B and C of the shelf, thus providing an important correlation marker that can be traced through the slope, reef, and shelf beds. The configuration of that correlation marker suggests that the Yates-equivalent Capitan reef had a very steep profile, similar to that of modern reefs. The Tansill-equivalent reef, well-exposed at Walnut Canyon, has been eroded at McKittrick Canyon; the only Tansill-equivalent reef present at McKittrick occurs as fallen blocks in the slope facies. Yates- and Tansill-equivalent reef deposits differ in that the older reef contains more internal sediment, less Archaeolithoporella/botryoidal-fan cement, and perhaps a greater variety of sponges.
| <<< Previous | <<< Contents >>> | Next >>> |
state/tx/1993-26/sec3.htm
Last Updated: 2-Apr-2007